Chromosomes et anomalies chromosomiques
Atlas of Genetics and Cytogenetics in Oncology and Haematology
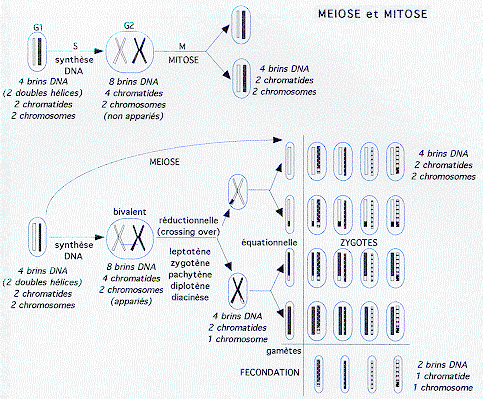
MITOSE / MÉIOSE
(Rappel)
CHROMOSOMES
INTRODUCTION
LES CHROMOSOMES sont le :
¤ support du matériel génétique
¤ support de l'hérédité
¤ support de l'organisation de la vie cellulaire
Hérédité: 1 chromosome maternel + 1 chromosome paternel (pour chaque paire de chromosomes homologues)
--> diploïdie (2 lots); copie exacte de l'ensemble passé ensuite à chaque cellule fille, de génération en génération cellulaire.
Vie cellulaire: qui peut être bouleversée
- soit lors de l'embryogénèse (anomalies constitutionnelles)
- soit lors d'un cancer (anomalies acquises)
NOMENCLATURE
Le caryotype humain normal comporte 46 chromosomes:
¤ 22 paires d'autosomes, notés de 1 à 22 en fonction de leur taille décroissante
¤ 1 paire de gonosomes, ou chromosomes sexuels: XX chez le sujet féminin, XY chez le sujet masculin.
Chaque chromosome comporte un centromère (CEN) région qui contient le kinetochore, centre d'organisation des microtubules responsable de la fixation des chromosomes au fuseau mitotique lors de la mitose. Les deux chromatides soeurs sont unies dans leur zone hétérochromatique de chaque coté du centromère.
Chaque bras se termine par un télomère (en pter et qter), séquence ADN répétitive hautement conservée qui empêche les fusions avec d'autres chromosomes. Rôle également de ces séquences dans l'attachement des télomères à l'enveloppe nucléaire, en particulier lors de la méiose. Le raccourcissement des télomères est un phénomène connu, et qui parait associé au vieillissement cellulaire.
De part et d'autre du centromère, une chromatide présente 2 bras: le bras court ou bras p, placé en haut sur un caryotype, et le bras long (bras q) placé en dessous du centromère.
Si le bras court est presque aussi long que le bras long, le chromosome est dit métacentrique; s'il est nettement plus court, le chromosome est dit sub-métacentrique. Quand il est encore plus court, le chromosome est appelé sub-télocentrique (le Y est sub-télocentrique). Enfin, si ce bras p est très petit, le chromosome est dit acrocentrique. Les acrocentriques sont les chromosomes 13, 14, 15, 21, 22. Les acrocentriques, portent sur leurs bras courts de l'hétérochromatine et des régions impliquées dans l'organisation nucléolaire (NOR) et codant pour l'ARN ribosomal.
Chaque bras est arbitrairement divisé en régions, notées de 1 jusqu'à 4 (pour certains chromosomes) en partant du centromère. Chaque région est divisée en bandes, entités visibles -pâles ou foncées- après usage d'une technique de dénaturation. Chaque bande peut, si nécessaire, être divisée en sous-bandes (chromosomes en prophase, moins condensés, et donc montrant plus de détails) ... Ainsi, un emplacement sera défini par le numéro du chromosome où se trouve cet emplacement, suivi de la lettre indiquant le bras impliqué, suivie des numéros de région, de bande, voire de sous-bande. Ex: l'emplacement noté d'une astérisque sur le schéma ci-contre sera localisé par la nomenclature en 21q22.3 (voir Figure).

ANOMALIES CHROMOSOMIQUES:
CONSTITUTIONNELLE/ACQUISE,
HOMOGÈNE/EN MOSAÏQUE,
DE NOMBRE/ DE STRUCTURE
* Une anomalie chromosomique peut être:
CONSTITUTIONNELLE:
les différents organes ("l'ensemble de l'individu") ont la même anomalie (voir Figure). L'accident chromosomique existait déjà chez l'embryon; il s'est produit avant la fécondation, dans l'un des gamètes, ou peu après, dans une des cellules du zygote. Le sujet porteur, si l'anomalie est non équilibrée (id est: si certains gènes sont présents en plus ou en moins de deux copies) a souvent une dysmorphie et/ou des malformations viscérales, et/ou un retard du développement psychomoteur (triade).
ACQUISE:
un seul organe est touché, les autres organes sont normaux. L'accident chromosomique s'est produit au cours de la vie de l'individu; il est acquis par rapport au caryotype constitutionnel. Le sujet est porteur d'un processus cancéreux sur l'organe impliqué.
* Une anomalie chromosomique peut être:
HOMOGÈNE:
si toutes les cellules du tissu examiné portent la même anomalie.
exemple 1: une anomalie constitutionnelle survenue chez un gamète parental (ex: + 21) se retrouvera chez toutes les cellules de l'enfant descendant (ex: trisomie 21 homogène).
exemple 2: une anomalie acquise survenue lors d'une leucémie peut être présente sur toutes les cellules sanguines étudiées chez cet individu (si les cellules sanguines normales sont suffisamment inhibées pour que l'on n'en retrouve aucune en mitose (ex: t(9;22) dans la leucémie myéloïde chronique (LMC)).
EN MOSAÏQUE:
si certaines cellules du tissu examiné portent l'anomalie alors que d'autres sont normales (notion de clone).
exemple 1: une anomalie constitutionnelle survenue chez le zygote après plusieurs divisions cellulaires (ex: +21) ne touchera qu'une partie des cellules de l'embryon puis de l'enfant (ex: 46, XY/47, XY, +21).
exemple 2: une anomalie acquise, dans une leucémie, peut n'être présente que sur une partie des mitoses si des cellules normales entrent en division; un clone supplémentaire peut porter des anomalies additionnelles (ex: 46, XY/46, XY, t(4;11)/46, XY, t(4;11) i(7) dans une leucémie aiguë lymphoblastique).
* Une anomalie chromosomique peut être
DE NOMBRE:
si un (des) chromosome(s) est (sont) surnuméraire(s) (trisomie) (ex: +21) ou manquant(s) (monosomie) ex: XO par perte d'un gonosome . Note: le caryotype est toujours déséquilibré lors d'une anomalie de nombre.
DE STRUCTURE:
si il y a cassures chromosomiques et recollements erronés ,l'anomalie de structure est:
¤ équilibrée: si il n'y a ni perte ni gain de matériel génétique
¤ déséquilibrée: si il en résulte une délétion et/ou une duplication d'un fragment.

ANOMALIES CHROMOSOMIQUES - MÉCANISMES ET NOMENCLATURE
I - ANOMALIE DU NOMBRE
A- HOMOGÈNES
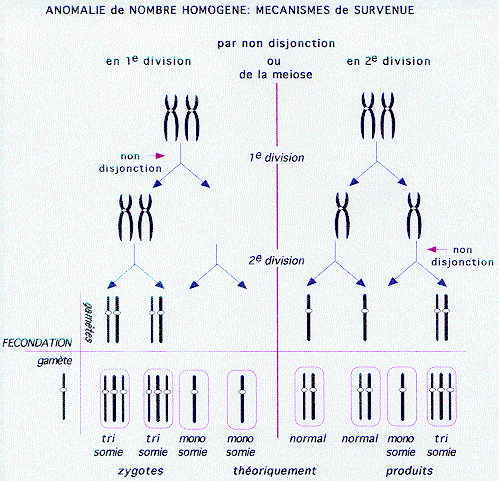
1 - Homogènes par non disjonction méiotique (voir Figure)
1.1. Autosomes
- une non disjonction en première division produit 4 gamètes déséquilibrés.
- une non disjonction en deuxième division produit 2 gamètes déséquilibrés et 2 gamètes normaux.
Les gamètes possédant un autosome en excès produisent un zygote trisomique: de nombreuses trisomies ne sont pas viables et involuent très précocement, ou sous forme de fausses couches spontanées (trisomie 16 par exemple). D'autres sont plus ou moins viables: trisomies 21, 13, 18, 8.
Les gamètes nullosomiques produisent des monosomies. La monosomie 21 serait, si elle s'avère exister, la seule monosomie d'autosome viable. Les monosomies, bien que produites en nombre théoriquement égal aux trisomies, subissent une élimination précoce encore plus stricte.
Les non disjonctions peuvent concerner chaque paire de chromosomes et peuvent être multiples au cours d'une méiose (les non disjonction multiples impliquent souvent un gonosome).
Il ne s'agit pas d'un événement rare, mais d'un phénomène très fréquent, compensé le plus souvent par l'élimination précoce du conceptus déséquilibré.
Chez l'homme les 4 types de gamètes représentés figurent effectivement dans le sperme. Chez la femme un seul participera à la fécondation, les trois autres étant des globules polaires, éliminés).
1.2. Gonosomes
Pour les gonosomes, la viabilité des conceptus déséquilibrés est plus grande, et le phénomène de non disjonction apparaît alors, dans la grande variété de ses conséquences.
Table: Formules gonosomiques des zygotes produits pour tous les types de gamètes obtenus. Les cases non remplies ne donnent pas de conceptus viables. Les cases XX et XY avec une ° sont des zygotes normaux issues de gamètes normaux. Les cases avec une * sont des zygotes normaux éventuellement produits par des gamètes dont les anomalies se sont compensées.
|
gamètes |
O |
Y |
X |
XY |
YY |
XX |
XYY |
XXY |
XXYY |
|
O |
|
|
X |
XY* |
|
XX* |
XYY |
XXY |
XXYY |
|
X |
X |
XY° |
XX° |
XXY |
XYY |
XXX |
XXYY |
XXXY |
|
|
XX |
XX* |
XXY |
XXX |
XXXY |
XXYY |
XXXX |
XXXXY |
|
|
|
XXX |
XXX |
XXXY |
XXXX |
XXXXY |
|
XXXXX |
|
|
|
|
XXXX |
XXXX |
XXXXY |
XXXXX |
|
|
|
|
|
|
2 - Homogènes par anomalie de la fécondation
Entraînent une polyploïdie
Triploïdies le plus souvent: 3N = 69 chromosomes 69, XXX ou 69, XXY ou 69, XYY. Présentes dans 20 % des fausses couches spontanées, elles peuvent aboutir à la naissance d'enfants vivants, qui meurent cependant très rapidement. Le mécanisme de formation des triploïdies est double:
1. la digynie: non expulsion du 2ème globule polaire.
2. la diandrie: fécondation d'un ovocyte I par 2 spermatozoïdes. La diandrie est 4 fois plus fréquente que la digynie.
Tétraploïdies: 4N = 92 chromosomes (très rares naissances vivantes décrites, rapidement fatales). Présentes dans 6 % des fausses couches spontanées.
Ces accidents de la fécondation sont donc banal et sont estimés à 2 à 3 % des oeufs fécondés.
B - MOSAIQUES
Un individu en mosaïque est constitué de deux (ou plus de deux) populations à contenu chromosomique différent, mais provenant du même zygote (une mosaïque est notée par une barre oblique entre les 2 clones décrits; ex: trisomie 21 en mosaïque 46, XY / 47, XY, +21).
Elle a pour cause une non-disjonction mitotique: 1 cellule fille emportera 3 chromatides sur les 4, l'autre cellule fille n'emportant qu'une chromatide sera donc monosomique pour ce chromosome. Les autre cellules présente chez ce zygote se divisent normalement. Dans l'exemple de la trisomie 21 en mosaïque ci-dessus, le clone monosomique 21, non viable a disparu.
L'intensité du phénotype dépend du dosage respectif de ces deux populations cellulaires.
Les mosaïques ont une composition variable en fonction:
¤ de la précocité de la (ou des) non disjonction(s) mitotique(s) (45, X / 47, XXX sans aucune cellule à 46: événement zygotique; 45, X / 46, XX / 47, XXX: événement post-zygotique, d'autant plus tardif que les cellules 46, XX sont plus nombreuses; si l'accident s'est produit au stade 32 ou 64 cellules, tout, ou partie, ou aucune cellule de l'embryon (ne) sera affecté, puisque les cellules du disque embryonnaire sont différenciées à ce stade, et que l'aberration touchera donc les annexes OU l'embryon.
¤ de la répartition des populations cellulaires au cours de l' embryogenèse. La proportion des différents clones peut varier d'un organe à l'autre.
¤ de la pression de sélection à laquelle ces populations peuvent réagir de façon différente, in vivo, avec avantage d'un clone sur un autre, ou combinaison chromosomique létale (in vitro également où les différents clones peuvent avoir une cinétique de croissance variable).
Un cas particulier de mosaïque est le monozygotisme hétérocaryote où deux jumeaux vrais sont par exemple l'un 46, XY, garçon normal, l'autre 45, X avec syndrome de Turner. Pour ce dernier, la perte de l'Y a coïncidé avec la séparation de l'oeuf pour former les deux jumeaux. La mosaïque s'est répartie sur deux personnes, qui ont le même matériel génétique à l'anomalie chromosomique près. Un certain nombre de cas ont été rapportés, avec des anomalies variées.
Une mosaïque ne doit pas être confondue avec une chimère qui est un organisme dont les cellules dérivent de deux zygotes (ou plus) distincts. Elles sont produites:
par mélange ou échange de cellules entre zygotes différents (fusion précoce de deux embryons, ou circulation croisée avec greffe des cellules échangées entre jumeaux dizygotes).
par syngamie: fécondation anormale avec, par exemple, dispermie, participation de l'ovotide et du deuxième globule polaire aboutissant à la constitution d'un individu XX/XY. (individu à "2 pères et 2 mères").
Dans les chimères, les caryotypes des différentes lignées peuvent être différents, et surtout les contenus géniques de ces lignées le sont toujours.
Note: Les mosaïques sont fréquentes dans les processus malins, soit parce que des cellules normales demeurent et entrent en division, soit parce que le clone malin engendre des sous clones porteurs d'anomalies additionnelles , dites aussi anomalies secondaires (notion d'évolution clonale). Au contraire, l'anomalie acquise est homogène, lorsque seules les cellules porteuses de l'anomalie ont pu être caryotypées. Ces cellules malignes ont envahi le tissu, il n'y a plus de cellules normales ou bien celles-ci n'ont pas été vues en mitose.
II - ANOMALIES DE STRUCTURE
(voir Figure; voir aussi l'article: An Introduction to Chromosomal Aberrations )
A - Introduction
Les chromosomes peuvent subir des cassures suivies de recollement des extrémités libres:
¤ ou bien le chromosome se recolle comme il était: reditus ad integrum
¤ ou bien existent plusieurs cassures et le recollement est erroné: apparition d'une aberration
L'apparition de cassures chromosomiques, compensées par l'action de systèmes de réparation est un phénomène cellulaire banal. On connaît des pathologies de ces systèmes de réparation chez l'homme (voir Cancers héréditaires ).
Le plus souvent le site de cassure ne se trouve pas dans une séquence codante et ne provoque pas de mutation.
Ces cassures peuvent affecter tous les chromosomes, y compris les bras courts des acrocentriques.
Ce qui compte pour un individu, c'est de posséder une double copie du message de l'espèce, pas plus et pas moins. Ceci est vrai pour l'embryon, où un déséquilibre de dosage génique entraîne un développement anormal, et donc une pathologie constitutionnelle (Note: en fait un équilibre génique total n'est pas nécessaire à certaines cellules différenciées, qui n'utilisent qu'une partie des fonctions protéiques disponibles au niveau génique. Cependant, de nombreux déséquilibres, qui ne touchent pourtant qu'un petit nombre de gènes, sont très délétères). Ceci est également vrai en génétique des processus malins où la perte (ou la mutation) d'un gène suppresseur du cancer (antioncogène) est un événement primordial (perte d'hétérozygotie; modèle rétinoblastome, voir Cancers héréditaires ): la cellule garde un comportement normal, mais la perte du deuxième allèle permet l'apparition de la tumeur.
Ce qui compte aussi, c'est qu'un recollement erroné de segment chromosomique n'inactive pas un gène important (ex où l'inactivation induit une maladie autosomale dominante) ni ne donne naissance à un néogène hybride qui coderait pour une protéine de fusion ayant des propriétés oncogéniques. (voir Hémopathies malignes).
Note: beaucoup d'anomalies structurales sont létales pour la cellule. Parmi celles qui ne le sont pas et sont transmises, les plus fréquentes sont les translocations, les délétions et les petites inversions.
Note: les chromosomes remaniés sont souvent appelés dérivés (der) et le dérivé porte le numéro du chromosome dont il possède le centromère.
B - Principales anomalies de structure (Figure)

1 - Translocation réciproque
Echange, entre 2 chromosomes, d'une partie d'un de leurs bras. Tous les chromosomes peuvent être concernés par un tel réarrangement.
Remaniement équilibré.
Noté t, suivi d'une parenthèse indiquant les numéros des 2 chromosomes impliqués, séparés d'un point virgule; une deuxième parenthèse indique les points de cassure sur chacun des 2 chromosomes.
Transmission à la descendance (génétique constitutionnelle)
En méiose, les chromosomes homologues s'apparient (en bivalent) et des échanges de chromatides peuvent se produire. En cas de translocation réciproque (voir Figure), la seule manière d'apparier chaque zone homologue est de former un tétravalent (ou quadrivalent). A la métaphase I de la méiose, il n'y a pas de clivage des centromères et le tétravalent se démantèle. Enfin , en anaphase II la ségrégation des chromatides peut se faire selon les modes suivants:
Le type alterné produit des gamètes normaux, ou porteurs équilibrés de la translocation parentale.
Le type adjacent 1 est fréquent: il associe un chromosome normal (ex; chromosome a) avec le remanié de l'autre paire (der(b)). Il y a alors "duplication-déficience": présence en excès d'une séquence et défaut d'une copie d'une autre séquence.
Le type adjacent 2 est très rare: il associe un chromosome normal avec son homologue remanié (ex: a + der(a)).
Le type 3:1 est rare: Soit un chromosome remanié est transmis avec les homologues normaux (ex: a, b, der(b)) dans une cellule fille, et l'autre remanié (der(a)) dans l'autre cellule; Soit un homologue normal est transmis avec les deux chromosomes remaniés (b, der(a), der(b)) dans une cellule, et l'autre chromosome normal (a) dans l'autre cellule). Contrairement aux cas précédents, les zygotes n'ont pas 46 chromosomes, mais 47 ou 45.
Caractéristiques:
Les translocations réciproques sont des remaniements équilibrés (lorsqu'ils se produisent), et le sujet porteur est de phénotype normal.
Ils exposent à des malségrégations (surtout s'il existe un acrocentrique dans la translocation), ce qui provoque alors un déséquilibre. D'une façon générale, les zygotes déséquilibrés produits sont d'autant moins viables que les segments qui ont été échangés sont plus grands.
Un échange de chromatides homologues en méiose n'aura pas de conséquence sur la structure des chromosomes impliqués.
Certaines translocations réciproques se font avec des points de cassure dans les centromères, et échanges complets de bras courts ou longs.
Translocations complexes:
Le même processus peut s'effectuer avec 3 (ou plus) chromosomes, et des transferts de segments en chaîne entre les chromosomes concernés. Les translocations équilibrés constatées sont généralement des translocations cycliques. Les techniques de FISH ont montrées que ces translocations complexes sont relativement fréquentes.
Note: les translocations réciproques peuvent intervenir à n'importe quel moment au cours de la vie; elles s'observent dans les processus cancéreux.

2 - Translocation Robertsonienne
Fusion de 2 acrocentriques par cassure-recollement à proximité des centromères, plus souvent sur les bras p, engendrant alors un chromosome dicentrique (dic), possédant 2 centromères. L'un des 2 centromères est généralement inactivé, permettant au chromosome remanié de se comporter comme un monocentrique, sans problème de ségrégation à l'anaphase. Le chromosome remanié comporte dans tous les cas les bras longs des 2 acrocentriques concernés, alors que les bras courts sont généralement perdus.
Le caryotype est pourtant dit équilibré, parce que la perte des bras courts de 2 acrocentriques est sans retentissement phénotypique. Le caryotype du porteur à phénotype normal comporte alors 45 chromosomes.
Noté t, suivi d'une parenthèse indiquant le numéro de chaque chromosome suivi de la notation q (ex: t(14q21q)).
Caractéristiques:
Les fusions centriques représentent l'anomalie chromosomique la plus fréquente dans la population humaine.
Le rôle physiologique des acrocentriques dans l'organisation nucléolaire rapproche leurs bras courts, et les expose à ce type de remaniement. Ces NORs actifs forment les nucléoles. Fréquemment, plusieurs de ces nucléoles fusionnent, mettant à proximité les uns des autres les bras courts des acrocentriques, ce qui favorisent les échanges, et donc les translocations. Au contraire, dans certains cas, la présence d'un nucléole peut être un barrière physique empêchant un rapprochement trop important et donc un risque de translocation.
Elles peuvent survenir de novo, ou être transmises.
Elles exposent à la constitution de gamètes déséquilibrés (voir Figure), graves pour les chromosomes 13 et 21 surtout, car viables en trisomie.
Toutes les combinaisons entre acrocentriques ne sont pas aussi fréquentes; la plus fréquente est la t(14q21q).
Les translocations entre homologues ne permettent de former que des gamètes déséquilibrés.
Les fusions centriques jouent un rôle important dans la spéciation.

3 - Délétion
Perte d'un segment au sein d'un chromosome (voir Figure); implique la perte des gènes portés par ce segment ("monosomie" partielle).
Remaniement déséquilibré.
Noté del, suivi du numéro du chromosome dans une première parenthèse, suivi dans une deuxième parenthèse des 2 points de cassure indiquant la région délétée (délétion interstitielle). Dans le cas où la délétion semble terminale, un seul point de cassure est noté. Cependant, un chromosome ne peut être stable, ont pensé les cytogénéticiens depuis longtemps, que s'il porte à chaque extrémité un télomère, structure particulière qui "ferme" le chromosome. Ainsi, toute délétion serait nécessairement de type interstitiel. Ceci a été récemment démontré par diverses techniques de cytogénétique moléculaire.
3.1. Délétion constitutionnelle
Sur un autosome, le retentissement phénotypique est majeur (exemples: del(18p) ; del(18q) ; del(4p): syndrome de Wolf ; del(5p): "cri du chat") Maladies chromosomiques constitutionnelles ); et le risque de transmission à une éventuelle descendance est donc exclu. Le remaniement survient donc de novo le plus souvent (10 à 15 % seulement des délétions proviennent de la malségrégation d'un remaniement parental équilibré; dans ce dernier cas, la délétion peut être pure, ou accompagnée d'une trisomie partielle pour un autre chromosome: (duplication/déficience); voir: translocations réciproques).
Cas particulier des microdélétions: elles sont parfois transmises (exemple: del(13)(q1400q1409): rétinoblastome).
Sur un gonosome, une délétion provoque des troubles de la différenciation sexuelle et de la gamétogénèse (sauf pour la partie distale de l'Yq) (ex: del(Xp): syndrome de Turner).
3.2. Délétion acquise: peut signifier la perte d'un gène suppresseur de cancer (exemple: del(13)(q14.00q14.09): rétinoblastome).

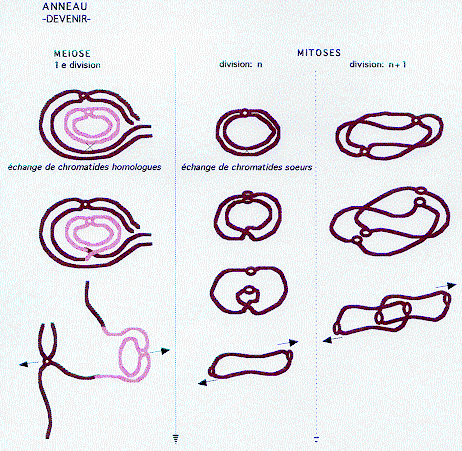
4 - Anneau
Délétions plus ou moins étendues (souvent peu importantes) aux deux extrémités d'un chromosome et recollement du segment intermédiaire (par absence de télomères), ce qui aboutit à une structure annulaire (il existe également des anneaux acentriques, mais ceux ci sont rapidement perdus, comme tout fragment ne possédant pas de centromère).
Remaniement déséquilibré.
Noté: r(ring), suivi du numéro du chromosome dans une parenthèse, suivi éventuellement d'une deuxième parenthèse indiquant les points de cassure, si ceux-ci sont localisables (ex: r(13)(p12q33)). Comme les anneaux se réarrangent sans cesse, les points de cassures varient, et leur notation n'est pas forcément pertinente.
Rarement transmis à la descendance (car un anneau est particulièrement instable; voir Figure), donc le plus souvent de novo.
Par échange de chromatides soeurs lors d'une mitose, l'anneau peut se dupliquer en anneau double dicentrique (voir Figure). Cet anneau entre alors dans le cycle fusion- fission. L'anneau peut alors être perdu (surtout si se forme un anneau acentrique), remanié (anneaux de tailles variables), en multiples copies, parfois intriquées.
--> Fréquence des mosaïques.
Phénotypiquement, le retentissement peut donc être celui d'une délétion, mais aussi d'une duplication.
Le plus fréquemment rencontré est l'anneau du 13.
5 - Inversion
Notée inv, suivi du numéro du chromosome dans une parenthèse, suivi d'une autre parenthèse indiquant les points de cassure. ex: inv(9) (p11q13).
Les inversions sont des remaniements équilibrés (quand elles se produisent); le porteur a un phénotype normal.
5a - Inversion paracentrique
Une inversion est paracentrique si un segment d'un bras de chromosome se casse (2 cassures sur un même bras), et se recolle tête bêche sur ce bras de chromosome.
Remaniement rare et/ou souvent non détecté (si seulement un très petit segment est inversé).
Les plus fréquemment rencontrées sont les inv para du 3, du 7 ou du 14.
Les sujets porteurs d'une inversion paracentrique sont souvent fertiles (les hommes autant que les femmes), leur descendance est pratiquement pour moitié de sujets normaux et pour l'autre moitié de sujets porteurs de l'anomalie équilibrée (il y a peu d'aneusomie de recombinaison, en partie parce que les gamètes déséquilibrés le sont généralement trop pour donner un conceptus). A noter quelques cas de descendants malformés apparemment équilibrés.
En méiose (voir Figure), les segments homologues des 2 chromosomes homologues s'apparient et forment une boucle d'inversion. Un échange de chromatides homologues dans la boucle d'inversion provoque le remaniement décrit en a: le fragment acentrique est perdu et se forme un pont chromosomique entre les 2 centromères attirés à chaque pôle. Ce pont:
1. casse: il y a alors, selon l'endroit de la cassure, duplication et déficience de certains segments dans la cellule fille, ou bien
2. empêche la diacinèse (séparation en 2 cellules filles): il en résulte 1 seule cellule fille portant le double de matériel génétique ; ou bien
3. le dicentrique sera exclu de chacune des 2 cellules filles (--> micronoyau); ou enfin
4. entraîne l'ensemble des deux homologues dans une seule cellule fille. Dans le dernier cas, on aura en télophase de deuxième division 1- 1 cellule normale (chromatide blanche sur le dessin) 2- 1 cellule porteuse de l'inversion équilibrée (chromatide noire) 3- 1 cellule ne portant aucun exemplaire de cette chromatide et 4- une cellule portant le dicentrique. Ce dicentrique va alors: soit i) entrer dans un cycle fission-fusion (remaniements complexes, nombreux), soit ii) empêcher la diacinèse provoquant une tétraploïdie, ou enfin iii) inactiver l'un de ses deux centromères et stabiliser le réarrangement.
D'autres échanges de chromatides homologues sont possibles, certains donnant naissance à 100 % de produits déséquilibrés (voir Figure).
En pratique, une forte pression sélective favorise les produits de méiose normaux ou portant l'inversion à l'état équilibré (comme souvent en génétique, une anomalie trop importante aura moins de répercussion qu'une anomalie moindre, par effet contre sélectif puissant; les gonies, oeufs, ou embryons porteurs seront rapidement éliminés: les échantillons observables sont hautement biaisés, ce qui est d'ailleurs une règle générale de la Biologie).).

5b - Inversion péricentrique
Une inversion est péricentrique si un chromosome se casse de part et d'autre du centromère (1 cassure sur chaque bras) et se recolle de manière erronée.
Certaines sont très fréquentes et on les inclut dans les variants chromosomiques plutôt que dans les anomalies:
inv(9)(p11q13): l'inversion péricentrique du chromosome 9 existe chez un sujet pour 400 environ (avec de grandes variations géographiques). On n'a pas décrit d'aneusomie de recombinaisons pour elle (les échanges de chromatides dans l'hétérochromatine sont exceptionnels), mais sa totale innocuité est encore controversée.
inv(Y): existe chez 1 à 2 sujets sur 1000 sujets masculins.
Une inversion péricentrique peut provoquer des fausses couches spontanées, parfois une stérilité chez l'homme, et des aneusomies de recombinaison à la méiose .
Un échange de chromatide homologue à la méiose, s'il se produit dans la boucle d'inversion, va faire apparaître des chromosomes recombinants (rec) portant une duplication d'un segment et une déficience d'un autre (une duplication de p - déficience en q sera notée rec dup(p)). Note: les segments dupliqués-déficients sont les segments se trouvant en dehors de la boucle d'inversion (voir Figure).
Si l'inversion est grande, le risque de crossing over dans la boucle d'inversion sera plus grand et les segments dupliqués-déficients (hors boucle) plus petits. Cependant, le risque est alors plus grand, car la possibilité que le conceptus soit viable est plus importante. Au contraire, une inversion petite implique un risque moindre d'échange de chromatides dans la petite boucle d'inversion. Si cet échange de chromatides intervient, les segments dupliqués-déficients, très importants auront un effet contre sélectif important, et le risque de conceptus malformé sera théoriquement moindre.
Note sur les inversions para et péricentriques:
Les échanges de chromatides survenant dans les zones non inversées sont sans conséquences.
Quel que soit le lieu de l'échange de chromatides dans la boucle, la conséquence est la même.
Deux, ou un nombre pair, d'échanges de chromatides dans la boucle s'annulent.

6 - Isochromosome
Perte d'un bras entier d'un chromosome, "remplacé" par la duplication de l'autre bras (donc équivalent d'une monosomie pour un bras et d'une trisomie pour l'autre: remaniement déséquilibré).
Remaniement déséquilibré.
Noté i suivi d'une parenthèse indiquant le numéro du chromosome impliqué et la lettre du bras dupliqué (ex: i(17q) ou i(17)(q10): perte du bras p et duplication du bras q).
Fréquence de ce réarrangement sur le chromosome X (syndrome de Turner par i(Xq)).
Fréquent en tant qu'anomalie acquise (exemple: i(17q), anomalie secondaire de la leucémie myéloïde chronique).
Les mécanismes de formation d'un isochromosome sont variés (voir Figure). Si il s'est formé en 1ère division de la méiose, le matériel génétique dupliqué sera hétérozygote; sinon (id est: formation en 2e division de méiose ou au cours d'une mitose), le matériel génétique dupliqué sera homozygote. Dans les cellules somatiques, le mécanisme de survenue le plus probable est la cassure/délétion de 2 chromatides soeurs dans une région peri centromérique avec recollement en U.

7 - Insertion
Un fragment de chromosome se casse, les régions flanquant ce fragment se recollent (comme dans une délétion), et le fragment se réinsère à un autre endroit, soit sur le même chromosome, soit sur un autre chromosome
Remaniement équilibré.
Noté ins, suivi d'une parenthèse indiquant le numéro du chromosome qui reçoit l'insertion, suivie éventuellement d'un point virgule indiquant le numéro du chromosome donneur du segment (si ce chromosome n'est pas le même que celui qui reçoit), suivi d'une deuxième parenthèse indiquant le point d'insertion suivi d'un point virgule seulement si le chromosome donneur est différent, suivi des points de cassure du fragment (ex: ins(2)(p13q31q34) et ins(5;2)(p12;q31q34): le fragment q31q34 d'un chromosome 2 est inséré respectivement en p13 du même chromosome 2, et sur un chromosome 5 en p12.
Une insertion peut être directe (dir ins) si le fragment garde son orientation par rapport au centromère (la bande la plus proximale restant la plus proche du centromère), (exemples ci-dessus: la bande q31 est notée avant la bande q34).
Une insertion peut être inversée (inv ins) si la bande la plus proximale du fragment se retrouve la plus éloignée du centromère (ex: ins(2)(p13q34q31) et ins(5;2)(p12;q34q31)).
Cette aberration peut rester équilibrée et stable dans les cellules somatiques au cours des générations cellulaires. Mais elle est très instable en méiose; dans beaucoup de cas, le segment inséré est trop petit pour induire la formation d'un tétravalent. Même ainsi, la ségrégation en première division de méiose implique que la moitié des gamètes soit déséquilibrée. Si au contraire ce segment est assez large pour permettre la formation occasionnelle d'un tétravalent, alors, comme le montre la Figure, 25% des gamètes seront normales si l'insertion est directe, 0% si elle est inversée (où il y a la difficulté supplémentaire d'un pont dicentrique et d'un fragment acentrique). (Note: il est difficile d'imaginer un appariement sans ³boucle², mais cette interprétation reste théorique. Il est possible que des complexes synaptonémaux au pachytène/diplotène montrant une telle configuration en boucle aient été vus, au moins chez la souris ou le hamster).
Petit exercice amusant pour les étudiants: trouver toutes les constitutions gamétiques résultant d'une insertion.

8 - Duplication
En tandem: un segment de chromosome se duplique in situ (1 ou plusieurs fois) sur ce chromosome, le fragment dupliqué gardant la même orientation que le fragment original.
En miroir: le fragment dupliqué prend une orientation inverse du fragment original.
Remaniement déséquilibré.
Noté dup, suivi du numéro du chromosome dans une première parenthèse, suivi dans une deuxième parenthèse des 2 points de cassure indiquant la région dupliquée.
9 - Dicentrique
Chromosome possédant deux centromères: issus de mécanismes variés tels que ceux précédemment décrits, c'est également et surtout le mode de recollement alternatif d'une translocation réciproque.
Remaniement déséquilibré.
Noté dic, ou psu dic (pseudo dicentrique), quand l'un des 2 centromères s'inactive.
Fréquent en cas de translocation robertsonienne; exceptionnel en génétique constitutionnelle en cas de translocation n'impliquant pas au moins un bras court d'acrocentrique. Rare en tant qu'anomalie acquise.
Un dicentrique (autre que t rob) est hautement instable à moins que:
l'un des centromères ne s'inactive.
LI>la distance intercentromérique soit très faible.
Les seules preuves de la présence de 2 centromères actifs sont:
la visualisation de ponts anaphasiques.
la constatation de non disjonctions.
la présence d'isochromosomes de chacun des deux chromosomes concernés, résultant de cassures du pont et de fusion latérale (type U: entre chromatides soeurs).
La preuve de l'inactivation d'un centromère est facilement apportée lorsque les 2 chromatides, au niveau de ce centromère, sont largement écartées en lieu de la constriction.
10 - Remaniements complexes
- Impliquant plus de 2 chromosomes et/ou plus de 3 points de cassures.
- Fréquence des porteurs malformés apparemment équilibrés dans la descendance (conseil génétique).
- Fréquent dans beaucoup de tumeurs solides et de lymphomes.
11 - Marqueur
Elément chromosomique non reconnaissable, noté Mar;
soit petit élément supplémentaire au caryotype constitutionnel, avec ou sans retentissement phénotypique (problème en diagnostic prénatal)
soit élément de taille variable, souvent importante, présent dans un processus cancéreux. Les techniques de FISH ont montré que ce sont des chromosomes hautement remaniés, comportant des segments de chromosomes variés.
12 - Fragments Minus; HSR
Fragments minus (notés DM: double minute); très petit(s) élément(s) supplémentaire(s), souvent par 2, acentriques; généralement très nombreux.
HSR: noté ainsi pour "homogeneously staining region": région de coloration homogène et de taille variable, souvent importante, présente au sein d'un ou de plusieurs chromosome(s). Expérimentalement, les HSR peuvent se rencontrer après une exposition chronique à certains toxiques.
DM et HSR sont des éléments hautement corrélés à la présence d'une amplification génique importante. Se rencontrent lors des processus malins, en particulier en cas de tumeur solide.
Contributors:
Jean-Loup Huret, Claude Léonard, John RK Savage.