
Evolution et génétique
STABILITE ET VARIABILITE DES GENOMES ET EVOLUTION
EXEMPLES DE RELATIONS ENTRE MÉCANISMES DE L'EVOLUTION ET GÉNÉTIQUE
Comment la transmission des innovations génétiques au sein des populations peut-elle être responsable de l'évolution des espèces?
1 La sélection naturelle, un moteur de l'évolution
1.1 INNOVATION GENETIQUE ET ENVIRONNEMENT: LE MELANISME INDUSTRIEL CHEZ LA PHALENE DU BOULEAU
1.1.1 Variation au sein d'une population:
La phalène du bouleau (Biston betularia) est un papillon, fréquent en Europe du nord, qui vole la nuit et se repose le jour dans les bouleaux. Cette espèce a été très étudiée depuis le XIXème siècle car elle présente deux formes principales: l'une claire (typica) et l'autre sombre ou mélanique (carbonaria), dont les fréquences ont considérablement varié dans les populations naturelles au cours des 150 dernières années.
Au milieu du XIXème siècle, la forme typica était largement majoritaire dans les populations anglaises. En 1848 on a capturé le premier individu carbonaria dans la région de Manchester. La fréquence de cette forme s'est alors accrue rapidement et les individus mélaniques ont ensuite été observés dans les autres régions industrielles de l'Angleterre.
On sait aujourd'hui que le déterminisme génétique de cette coloration est monogénique et autosomique, l'allèle carbonaria (C) étant dominant sur l'allèle typica (c):
|
Formes mélaniques |
Forme claire | |
|
C / C |
C / c |
c / c |
|
|
|
|
Depuis 1848, les variations s'étant toujours produites dans le sens de l'augmentation de la fréquence de la forme carbonaria, il ne peut s'agir d'un phénomène dû au hasard. Essayons d'en comprendre les mécanismes.
1.1.2 L'hypothèse de mutations récurrentes:
L'allèle carbonaria pourrait être obtenu par mutation et remplacer l'allèle typica. On peut alors calculer le taux de mutation de c vers C, appelons-le v.
Appelons q la fréquence de l'allèle récessif c. Entre deux générations successives to et t1, la fréquence q1 devient:
q1 = q0 - vq0
En effet, la fréquence des allèles c sera celle de la génération précédente (q0) diminuée de la fréquence des allèles mutés (vq0). On considère ici que l'allèle C ne mute pas vers c, ce qui correspond à l'évolution la plus rapide possible sous l'effet de la mutation.
Le processus se répétant dans le temps, au cours des générations, on en déduit:
![]()
ou t représente le nombre de générations. Déterminons le taux de mutation v:
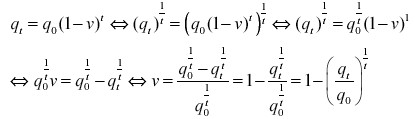
Nous avons donc:
![]()
![]()
![]() Si on considère que la fréquence phénotypique de la forme mélanique était de 0,01 dans la région de Manchester en 1848 et de 0,98 en 1895, et sachant que la phalène du bouleau présente une génération par an dans cette région, calculez la valeur de v qui rend le mieux compte de la vitesse de l'évolution observée?
Si on considère que la fréquence phénotypique de la forme mélanique était de 0,01 dans la région de Manchester en 1848 et de 0,98 en 1895, et sachant que la phalène du bouleau présente une génération par an dans cette région, calculez la valeur de v qui rend le mieux compte de la vitesse de l'évolution observée?
N.B. Le calcul (hors programme) des fréquences alléliques de la forme claire pour les deux valeurs ci-dessus donne respectivement 0,995 et 0,141.
Appliquez la relation permettant de calculer le taux de mutation v. Quelle remarque vous suggère le résultat obtenu? Peut-on valider l'hypothèse envisagée?
1.1.3 L'hypothèse sélective:
Dès le XIXème siècle, on remarqua une corrélation positive entre la fréquence de la forme mélanique et la pollution industrielle. Celle-ci contribue à la disparition des lichens sur les bouleaux et au noircissement de ces arbres. Le phénomène fut décrit sous le terme de "mélanisme industriel".
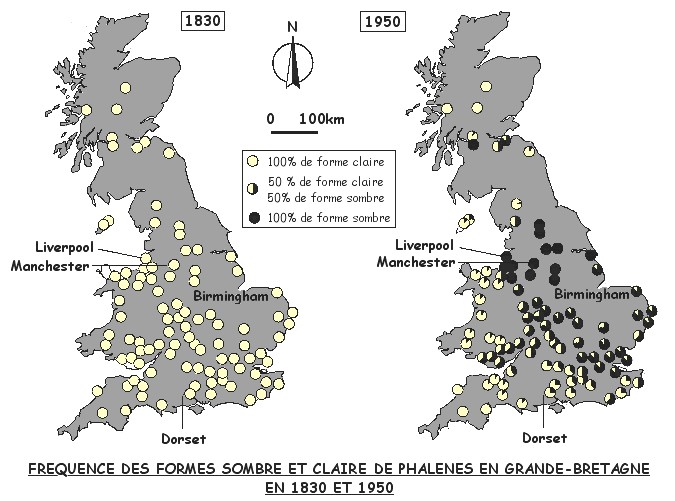
La distribution géographique des différentes formes est très suggestive: Les deux cartes ci-dessus présentent la distribution des fréquences des deux formes vers 1830 et 1950.
![]()
![]() Que constatez-vous?
Que constatez-vous?
Le rejet du modèle lamarckien
L'hypothèse la plus répandue supposait une action dirigée (non aléatoire) de l'environnement, modifiant un caractère héréditaire d'un organisme pour mieux adapter celui-ci à son milieu. Le caractère serait alors transmis sous la forme nouvelle. Cette hypothèse a été formalisée par LAMARCK (1744-1829). Elle est souvent présentée sous le terme d'hérédité des caractères acquis.
Compte tenu des connaissances actuelles sur le matériel génétique cette hypothèse est difficilement concevable. Il faudrait en effet que l'environnement "reconnaisse" le gène adéquat pour le modifier spécifiquement dans le "bon sens".
Le modèle darwinien
Dès 1897, certains chercheurs proposèrent une explication au mélanisme industriel dans le cadre de la sélection naturelle: on suggéra que les individus typica, posés sur des bouleaux clairs couverts de lichens, étaient mieux camouflés des oiseaux prédateurs que les individus carbonaria. Sur des bouleaux noircis par les suies et dépourvus de lichens, c'est l'inverse qui se produisait. Les deux formes seraient alors consommées de façon différente selon le type de support, la forme mélanique étant avantagée sur des arbres noircis. Elle se serait ainsi répandue facilement dans les régions industrialisées, suite à la modification de la surface des bouleaux par la pollution.
Cette hypothèse fut reprise par de nombreux biologistes et statisticiens et donna l'objet de simulations mathématiques qui contribuèrent à la théorie synthétique de l'évolution. Nous en reprendrons quelques aspects avec modestie dans le paragraphe suivant.
1.2 LA SELECTION NATURELLE: UNE REPRODUCTION DIFFERENTIELLE DES INDIVIDUS ET DES GENES:
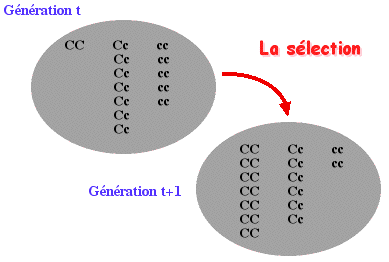 Dans la nature, les individus sont soumis à différentes conditions environnementales qui modulent le nombre de leurs descendants. De nombreux caractères sont impliqués dans ce processus. Pour les caractères possédant un déterminisme génétique, différents phénotypes (correspondant à différents génotypes) sont présents dans la population.
Dans la nature, les individus sont soumis à différentes conditions environnementales qui modulent le nombre de leurs descendants. De nombreux caractères sont impliqués dans ce processus. Pour les caractères possédant un déterminisme génétique, différents phénotypes (correspondant à différents génotypes) sont présents dans la population.
Dans un environnement donné, si les individus porteurs de ces phénotypes se reproduisent mieux que d'autres, ils auront tendance à transmettre plus fréquemment les gènes dont ils sont porteurs .
C'est l'idée de base présentée par DARWIN et WALLACE (1858), qui proposent que les facteurs héréditaires avantageant certains individus soient ainsi mieux transmis au cours des générations. Ils associent l'idée d'adaptation à celle de plus forte reproduction. Il s'agit là pour eux du processus fondamental des changements évolutifs, qu'ils appellent sélection naturelle. En termes actuels, il s'agit d'une transmission différentielle des gènes, et la "survie du plus apte" concerne en fait la capacité à transmettre plus efficacement certains types de gènes.
1.2.1 La modélisation
De nombreux travaux ont cherché à modéliser les effets de la sélection naturelle sur la variation des fréquences alléliques, au cours des générations. Le paramètre de base permettant de quantifier l'action de la sélection est appelé valeur sélective ou valeur " adaptative " du phénotype ("Darwinian fitness"). Il est classiquement noté w.
Dans la pratique, phénotype et génotype sont liés par les règles du déterminisme génétique et l'on associe directement au génotype la valeur sélective du phénotype qu'il détermine. Nous parlerons donc également des valeurs sélectives des différents génotypes, soit dans le cas d'un locus autosomique diallélique comme celui qui concerne la variété "typica" (c) et "carbonaria" (C):
|
Valeurs sélectives |
w11 |
w12 |
w22 |
|
Génotypes [phénotypes] |
CC [C] |
Cc [C] |
cc [c] |
De très nombreux composants peuvent contribuer à la valeur sélective d'un individu: elle dépend de la probabilité de survie jusqu'à l'âge de la reproduction, du génotype considéré et de sa fécondité. Les valeurs sélectives peuvent être estimées par le nombre moyen de descendants (viables et fertiles) fourni par un individu porteur d'un génotype, à la génération suivante. Dans la pratique on rapporte les valeurs absolues à la valeur du génotype le "meilleur" (valeur sélective relative).
Ainsi dans le cas théorique ci-dessous, voilà comment estimer les valeurs sélectives:
|
Génotypes |
CC |
Cc |
cc |
|
Nombre d'individus viables à la génération t |
30 |
50 |
20 |
|
Nombre d'individus viables fournis à la génération t+1 |
60 |
90 |
30 |
|
Valeurs sélectives absolues |
60 / 30 = 2 |
90 / 50 = 1,8 |
30 / 20 = 1,5 |
|
Valeurs sélectives relatives |
w11 |
w12 |
w22 |
|
|
2 / 2 = 1 |
1,8 / 2 = 0,9 |
1,5 / 2 = 0,75 |
![]()
![]()
![]() On peut estimer que les valeurs sélectives dans l'exemple du mélanisme sont les mêmes pour les génotypes CC et Cc. On supposera que la fréquence de l'allèle C soit passée de 0,005 en 1848 à 0,86 en 1895, soit en une cinquantaine de générations.
On peut estimer que les valeurs sélectives dans l'exemple du mélanisme sont les mêmes pour les génotypes CC et Cc. On supposera que la fréquence de l'allèle C soit passée de 0,005 en 1848 à 0,86 en 1895, soit en une cinquantaine de générations.
En utilisant le programme informatique Natural Selection "Kent HOLSINGER" de l'Université du Connecticut, réalisez un certain nombre de simulations:
1 - Sélectionnez une fréquence allélique p de départ parmi celles proposées. Le nombre de générations est fixé à 100.
2 - Sélectionnez une valeur sélective à chacun des trois génotypes comme le suggère le tableau ci-dessous:
|
w11 |
w12 |
w22 |
|
1,2 |
1,2 |
0,8 |
|
1,2 |
1,2 |
1,1 |
|
1,2 |
1,2 |
1,2 |
3 - Cliquez sur "start" pour commencer une simulation (vous pouvez cumuler et superposer les 3 simulations).
D'après vos résultats quelles sont les valeurs de w qui se rapprochent le plus de l'évolution des fréquences des allèles de coloration observée chez la Phalène du bouleau entre 1848 et 1895? Justifiez-votre choix.
1.2.2 Expériences de captures et recaptures
L'hypothèse sélectionniste fut développée par différents chercheurs et mise à l'épreuve dans les années 1950. On utilisa la méthode des captures-recaptures: un grand nombre de phalènes des deux formes furent marquées ventralement, puis relâchées. Quelques jours après, une nouvelle campagne de captures fut menée, et les individus marqués et non-marqués des deux formes furent dénombrés.
Sont présentés ici les résultats d'expériences de captures-recaptures effectuées dans deux régions proches, mais différentes par leur taux de pollution. Le tableau suivant résume les résultats obtenus dans des zones boisées "non polluées" de la région du Dorset et dans celles "polluées" situées à proximité de Birmingham (KETTLEWELL, 1955):
|
Dorset 1955 |
Forme [carbonaria C] |
Forme [typica c] |
Total |
|
Nombre d'individus marqués relâchés |
473 |
496 |
969 |
|
Nombre d'individus marqués recapturés |
30 |
62 |
92 |
|
% d'individus marqués recapturés |
|
|
|
|
Birmingham 1955 |
Forme [carbonaria C] |
Forme [typica c] |
Total |
|
Nombre d'individus marqués relâchés |
154 |
64 |
218 |
|
Nombre d'individus marqués recapturés |
82 |
16 |
98 |
|
% d'individus marqués recapturés |
|
|
|
![]()
![]() Evaluez puis analysez les différents taux de recapture entre les régions: y-a-t'il des différences statistiquement significatives pour les taux de recapture, en fonction des régions?
Evaluez puis analysez les différents taux de recapture entre les régions: y-a-t'il des différences statistiquement significatives pour les taux de recapture, en fonction des régions?
La technique de capture-recapture peut-elle être responsable des différences observées?
KETTLEWELL interprète ces données en terme de survie différentielle.
![]()
![]() Vous pouvez alors en déduire une estimation des valeurs sélectives, en considérant les taux de recapture comme représentatifs des taux de survie:
Vous pouvez alors en déduire une estimation des valeurs sélectives, en considérant les taux de recapture comme représentatifs des taux de survie:
|
|
Dorset 1955 |
Birmingham 1955 | ||
|
|
Forme mélanique CC et Cc |
Forme claire cc |
Forme mélanique CC et Cc |
Forme claire cc |
|
Taux de survie ou valeur sélective absolue |
|
|
|
|
|
Valeur sélective relative |
w11 ou w12 |
w22 |
w11 ou w12 |
w22 |
|
|
|
|
|
|
Après avoir calculé les valeurs sélectives relatives à partir des taux de survie, vous choisirez parmi les hypothèses ci-dessous celles qui vous paraissent convenir au cas étudié:
1 - les formes claires sont désavantagées dans les régions polluées et inversement,
2 - les formes mélaniques sont avantagées dans les régions non polluées et inversement,
3 - aucune des formes ne présente d'avantage relatif,
4 - les formes mélaniques sont avantagées dans les régions polluées et inversement.
1.2.3 Le rôle joué par les prédateurs: la notion de contrainte sélective
L'interprétation générale est que la variation de fréquence des formes de la phalène est due à la prédation différentielle des oiseaux selon les régions, et aux migrations des papillons entre régions.
Pour vérifier la contrainte sélective portée sur certains papillons dans un environnement donné, KETTLEWELL a exposé à la prédation par les oiseaux le même nombre de papillons des deux formes sur chacun des deux sites et a compté le nombre de papillons capturés par les oiseaux dans chaque cas:
|
Dorset 1955 |
Forme [carbonaria C] |
Forme [typica c] |
|
Nombre de papillons exposés aux oiseaux |
190 |
190 |
|
Nombre de papillons capturés par les oiseaux |
164 |
26 |
|
% d'individus capturés par les oiseaux |
|
|
|
Birmingham 1955 |
Forme [carbonaria C] |
Forme [typica c] |
|
Nombre de papillons exposés aux oiseaux |
58 |
58 |
|
Nombre de papillons capturés par les oiseaux |
15 |
43 |
|
% d'individus capturés par les oiseaux |
|
|
![]()
![]() Montrez que l'évolution des fréquences alléliques résulte bien d'un processus de sélection naturelle et justifiez qu'une innovation puisse se révéler tantôt favorable, tantôt défavorable selon les contraintes sélectives.
Montrez que l'évolution des fréquences alléliques résulte bien d'un processus de sélection naturelle et justifiez qu'une innovation puisse se révéler tantôt favorable, tantôt défavorable selon les contraintes sélectives.
1.2.4 Contre-épreuve : les conséquences de la dépollution
 L'interprétation sélectionniste proposée a pu être mise à l'épreuve de façon inattendue. En effet, dans les années 1950, la Grande-Bretagne adopta une législation anti-pollution ("the Clean Air Acts") qui eut pour effet de réduire les émissions de suie et de SO2.
L'interprétation sélectionniste proposée a pu être mise à l'épreuve de façon inattendue. En effet, dans les années 1950, la Grande-Bretagne adopta une législation anti-pollution ("the Clean Air Acts") qui eut pour effet de réduire les émissions de suie et de SO2.
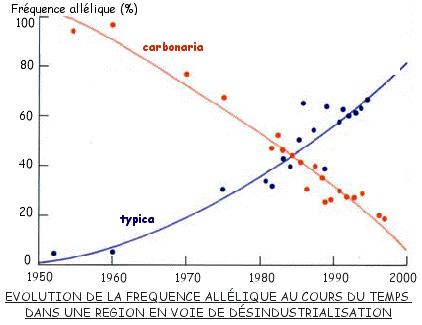
Dans la période qui suivit on observa une diminution de la fréquence de la forme mélanique, qui se poursuit depuis lors.
Le graphe ci-contre présente les observations effectuées dans une banlieue en voie de désindustrialisation.
1.2.5 Conclusion: la notion d'avantage sélectif
Selon l'environnement, l'une des formes de papillons, mieux "camouflée", échappe à ses prédateurs. Sa contribution à la formation de la génération suivante est proportionnellement plus importante: ainsi la fréquence des allèles qu'elle possède augmente dans la population et l'une des formes devient rapidement majoritaire. C'est la sélection naturelle: une mutation, phénomène aléatoire et "non dirigé", confère aux individus qui en sont porteurs un avantage sélectif; ceux-ci se répandent dans la population. Inversement les mutations défavorables ont tendance à être éliminées, car les individus qui en héritent survivent plus difficilement et se reproduisent moins. Ainsi, la sélection naturelle apparaît-elle comme un phénomène "dirigé".
Dans l'exemple choisi, tous les mélanismes observés ne sont pas de type "industriel". Il convient donc de se rappeler que les facteurs de la sélection naturelle peuvent être multiples et que chaque cas doit être étudié de manière approfondie en évitant les généralisations trop hâtives.
La sélection naturelle est relative à des conditions environnementales données. Un allèle n'est pas avantageux par essence, mais il le devient dans un environnement donné et cet état est susceptible d'évoluer.
1.3 FREQUENCE D'UN ALLELE ET CONTRAINTES SELECTIVES: LA DREPANOCYTOSE
Des exemples montrent qu'un allèle peut être soumis à des contraintes sélectives multiples, opposées et simultanées: l'exemple de la drépanocytose et du paludisme en est un.
L'allèle HbS présente un désavantage à l'état homozygote: il se traduit par une maladie grave de l'hémoglobine (la drépanocytose ou sicklémie).
Ce même allèle confère aux hétérozygotes, non atteints de la maladie, une résistance plus grande au paludisme, sévissant dans les mêmes régions que la sicklémie.
Il en résulte que la sélection naturelle maintient dans les populations un équilibre de la fréquence des allèles du gène: on parle de polymorphisme équilibré.
2 Variabilité génétique sans avantage sélectif
2.1 VARIABILITE ET FONCTIONS DE LA MYOGLOBINE
 La myoglobine est une protéine, présente chez tous les Vertébrés, dont la fonction est de fixer le gaz dioxygène dans le muscle. Une étude de la molécule avec un logiciel en 3D (Rasmol) révèle la relation entre structure et fonction: la chaîne polypeptidique forme, en un endroit, une "poche" dans laquelle un groupement non protéique appelé "hème" comporte un atome de fer capable de fixer 2 atomes d'oxygène.
La myoglobine est une protéine, présente chez tous les Vertébrés, dont la fonction est de fixer le gaz dioxygène dans le muscle. Une étude de la molécule avec un logiciel en 3D (Rasmol) révèle la relation entre structure et fonction: la chaîne polypeptidique forme, en un endroit, une "poche" dans laquelle un groupement non protéique appelé "hème" comporte un atome de fer capable de fixer 2 atomes d'oxygène.
Sur l'image ci-contre, quelques acides aminés désignés par une lettre et la position dans la chaîne ont été repérés.
![]()
![]() A l'aide du logiciel PHYLOGENE, vous afficherez les séquences des globines de quelques Vertébrés.
A l'aide du logiciel PHYLOGENE, vous afficherez les séquences des globines de quelques Vertébrés.
Pourquoi peut-on dire que certains acides aminés sont plus impliqués que d'autres dans la fonction de la molécule? Donnez quelques exemples au vu de l'image ci-contre.
Que constatez-vous chez la plupart des Vertébrés concernant ces acides aminés?
Etant donné le caractère aléatoire des mutations peut-on considérer que les portions d'ADN codant pour ces acides aminés ont échappé aux mutations?
Quelle a pu être, à leur égard, les effets de la sélection naturelle?

2.2 LES MUTATIONS NEUTRES
Dans d'autres régions de la molécule de myoglobine, sans relation directe avec sa fonction, on constate une grande différence d'une espèce à l'autre. De telles mutations ne modifient pas l'activité de la molécule et ne procurent ni avantage, ni désavantage sélectif aux individus qui en héritent: on dit que ces mutations sont neutres.
Au vu de ce que l'on a observé avec le mélanisme industriel, on pourrait penser à priori que la fréquence d'un allèle qui ne confère aucun avantage, ni désavantage sélectif reste stable (hypothèse).
![]()
![]()
![]() En utilisant le programme informatique Genetic Drift "Kent HOLSINGER" de l'Université du Connecticut, réalisez huit simulations:
En utilisant le programme informatique Genetic Drift "Kent HOLSINGER" de l'Université du Connecticut, réalisez huit simulations:
1 - Sélectionnez une fréquence allélique p de départ: 0,5. L'effectif de la population est restreint (au plus 250). Le nombre de générations est fixé à 100 par défaut.
2- Cliquez sur "start" pour commencer une simulation (vous pouvez cumuler et superposer les 8 simulations). Faites un essai de 8 simulations pour une population très restreinte, un autre pour une population plus grande.
Que constatez-vous? L'hypothèse émise ci-dessus est-elle vérifiée?
Mathématiquement l'hypothèse pourrait s'avérer exacte si l'effectif des populations était infini. En réalité la fréquence d'un allèle sélectivement neutre évolue de façon aléatoire d'autant plus que la population est restreinte: ce phénomène est qualifié de dérive génétique et signifie qu'une mutation neutre peut tout aussi bien se répandre dans une population ou être éliminée.
Son évolution est indépendante des conditions environnementales: en effet ceci est étayé par le fait que la comparaison des séquences de deux chaînes de globines, chez l'homme ou entre l'homme et une autre espèce vivant dans un environnement différent, montre un nombre de mutations identique: elles se sont accumulées à la même vitesse dans des milieux où les contraintes sont différentes.
Dans la dérive génétique, l'un des allèles dérive jusqu'à la fréquence 1, l'autre est perdu. La dérive vers l'homozygotie est plus rapide dans une population petite, mais l'issue est la même.
3 Chronologie du développement et évolution
Nous aborderons le sujet, difficile et relativement récent, de la façon suivante: les données anatomiques, chromosomiques et moléculaires ont permis de montrer, dans le chapitre "Parentés entre êtres vivants ...", que la proximité génétique de l'Homme et des Chimpanzés était très grande (génomes très probablement communs à 99%).
Il pourrait paraître paradoxal que des différences génétiques quantitativement si faibles aient des répercussions aussi fortes. N'est-ce pas parce que l'expression de certains gènes a des retentissements considérables au niveau du phénotype?
3.1 DEVELOPPEMENT DE L'HOMME ET DU CHIMPANZE
La tête et le crâne du jeune chimpanzé sont très différents de ceux de l'adulte et ressemblent beaucoup à ceux de l'homme.
![]()
![]() Vous analyserez les documents "Chronologies comparées du développement du chimpanzé et de l'Homme" ( J. Aubry - Lycée Charles de Gaulle - CAEN ) et vous essaierez de répondre au questionnaire présenté.
Vous analyserez les documents "Chronologies comparées du développement du chimpanzé et de l'Homme" ( J. Aubry - Lycée Charles de Gaulle - CAEN ) et vous essaierez de répondre au questionnaire présenté.
3.2 LES GENES DU DEVELOPPEMENT
3.2.1 La découverte des gènes du développement
Le programme de développement d'un organisme est inscrit dans ses gènes. Les cellules issues d'une même cellule-oeuf ayant la même information génétique mais des destinées différentes, on s'est longtemps demandé ce qui contribuait à la position des différents organes au cours du développement.
Les premiers éléments de réponse ont été fournis par l'étude de certaines mutations génétiques changeant le cours du développement chez la drosophile..." Anne Florimond - INRP - D'après "De la mouche à l'homme : un même supergène pour l'oeil". La Recherche, Février 1995.
3.2.2 Disposition et expression spatio-temporelle des gènes homéotiques chez la drosophile et la souris
"[...] l'ordre d'expression normal des gènes homéotiques le long de l'axe antéro-postérieur du corps correspond à leur ordre sur les chromosomes et [...] il est identique chez la Drosophile et chez l'embryon de souris" - INRP.
Nous retiendrons donc, comme le suggèrent les documents d'accompagnement du programme de SVT de terminale S, "qu'une mutation peut affecter des gènes du développement (notamment des gènes homéotiques). De telles mutations (« petite cause ») peuvent induire des décalages (ralentissements ou accélérations) de certaines phases du développement chez de nombreux vertébrés et invertébrés, et peuvent avoir par exemple pour conséquence l'apparition d'un nouveau plan d'organisation [...] (« grand effet »). On appelle hétérochronie ces modifications de durée et de vitesse d'ontogenèse au cours de l'évolution.
Ce mécanisme, qui peut contrôler l'apparition de caractères fondamentaux de l'espèce, peut être expliqué avec l'exemple du blocage du trou occipital à la base du crâne chez l'Homme qui impose une bipédie permanente, alors que la bascule vers l'arrière chez le Chimpanzé âgé de un an accompagne la quadrupédie des adultes.
Il en est de même avec l'exemple de l'allongement de la durée du développement embryonnaire du système nerveux central chez l'Homme par rapport au Chimpanzé, pouvant être lié à une multiplication des cellules nerveuses conférant à l'Homme un développement plus important du cerveau.
Les phénomènes d'hétérochronie ont probablement joué un grand rôle dans l'évolution humaine. Ils se manifestent par une accélération ou un retard dans le développement embryonnaire ou dans la croissance. Ainsi, l'Homme pourrait être le " descendant néoténique " d'un animal qui aurait conservé à l'âge adulte des traits que l'on trouve aussi chez les homininés actuels (Chimpanzé, Bonobo), mais seulement lorsque ceux-ci sont jeunes.
Ces caractéristiques sont modifiées lors du passage à l'état adulte (bipédie, front haut et bombé, yeux volumineux par rapport au reste de la face, membres supérieurs courts par rapport aux membres inférieurs). Des mutations finalement assez limitées sur le plan de la quantité de matériel génétique concernée, mais ayant des conséquences majeures sur le développement, ont pu être transmises d'un coup à la descendance."