
Système nerveux
FONCTIONNEMENT DU SYSTÈME NERVEUX
Nous allons acquérir les notions de base sur la communication nerveuse, puis élargir la compréhension des relations entre le phénotype et le génotype, pour étudier la part de l'expérience individuelle et celle de l'hérédité dans le remodelage permanent des circuits nerveux.
Eléments requis:
L'intégration des fonctions dans l'organisme
Thèmes traités et activités pédagogiques (durée: 6 semaines):
![]() Les circuits neuroniques d'un réflexe
Les circuits neuroniques d'un réflexe![]() Les messages nerveux
Les messages nerveux![]() Plasticité du système nerveux central
Plasticité du système nerveux central
Thème 1: Les circuits neuroniques d'un réflexe
La réalisation d'un mouvement nécessite la contraction de muscles précis. Commandée par le système nerveux, elle peut correspondre:
soit à un mouvement volontaire (activité spontanée),
soit à un mouvement automatique, involontaire en réaction à une stimulation (activité réflexe).
1.1 LE REFLEXE MYOTATIQUE:
Un réflexe se définit comme une réponse d'origine nerveuse, indépendante de la volonté, automatique, en réplique à une stimulation. Dans le cas d'un réflexe myotatique, il s'agit de la contraction réflexe d'un muscle déclenchée par son propre étirement. De tels réflexes interviennent dans le maintien de la posture pour s'opposer aux effets de la gravité. Ils sont testés par le médecin (réflexe rotulien, réflexe achilléen). Ce dernier, par exemple est déclenché par un choc bref sur le tendon d'Achille et se manifeste par une extension du pied, conséquence de la contraction du triceps sural du mollet.
1.1.1 Enregistrement du réflexe achilléen par ExAO
Principe de l'enregistrement: quand un muscle se contracte, il est le siège d'une activité électrique. A l'aide d'électrodes réceptrices et d'un amplificateur (module électrophysiologique), il est possible d'enregistrer cette activité: c'est l'électromyographie. Les tracés obtenus (électromyogrammes) traduisent ainsi l'activité électrique musculaire.
Protocole: trois électrodes sont placées sur le muscle du mollet (triceps sural = extenseur du pied) comme l'indique le dessin ci-contre à droite.
Le réflexe est déclenché en frappant à l'aide du marteau qui est connecté au dispositif d'ExAO (schéma ci-contre à gauche). Au moment du choc sur le tendon d'Achille, un contact électrique déclenche un signal qui détermine l'enregistrement des données fournies par les électrodes et leur traitement par le biais du logiciel installé sur l'ordinateur. En même temps, cela provoque la contraction réflexe du muscle extenseur du pied.
Enregistrement:
![]()

![]() Vous réaliserez cette expérience et vous imprimerez l'enregistrement obtenu. En voici un exmplaire ci-dessous:
Vous réaliserez cette expérience et vous imprimerez l'enregistrement obtenu. En voici un exmplaire ci-dessous:
Repérez sur l'enregistrement le délai qui existe entre l'instant du choc et celui de la réponse musculaire. Sachant que la contraction musculaire est déclenchée par des messages nerveux en provenance de la moelle (et non directement par l'étirement du tendon) déterminez le trajet du message nerveux responsable de la réponse réflexe. Evaluez-le et donnez ainsi un ordre de grandeur de la vitesse du message nerveux.
Si vous cherchez à modifier l'intensité du stimulus porté sur le tendon d'Achille, que révèle l'intensité de la réponse réflexe?
1.1.2 Des éléments anatomiques et histologiques interviennent dans la mise en oeuvre du réflexe achilléen. Nous les étudierons dans le cadre d'un T.P. d'histologie, leur concours à la réalisation du réflexe sera justifiée juste après. Ce sont:
des mécano-récepteurs, ici des fuseaux neuro-musculaires, stimulés par l'étirement: éléments constitués d'une capsule fibreuse contenant des fibres musculaires entourées des terminaisons de fibres ( ces dernières sont dénommées Ia sur la figure que vous découvrirez en cliquant sur l'icône ci-contre->). Ils sont placés en parallèle des fibres musculaires. Les fuseaux neuro-musculaires détectent la longueur des muscles et leur tension et transmettent cette information par les fibres afférentes* Ia.
* on qualifie d'afférentes les fibres nerveuses qui partent des récepteurs sensoriels, tels que les fuseaux neuromusculaires et qui arrivent au centre nerveux comme la moelle épinière. Par opposition, on qualifie d'efférentes les fibres nerveuses qui partent du centre nerveux pour se diriger vers les effecteurs périphériques, tels que les muscles.
des fibres nerveuses sensitives, logées dans le nerf rachidien sciatique, qui conduisent à la moelle les messages afférents ou centripètes,
des fibres nerveuses motrices du nerf sciatique, transmettant les messages efférents ou centrifuges aux effecteurs "muscles".
![]()
![]()
![]()
![]() Vous observerez deux préparations de nerf rachidien du commerce:
Vous observerez deux préparations de nerf rachidien du commerce:
- une coupe transversale de nerf rachidien, tel que le sciatique qui innerve le membre inférieur (ou postérieur chez les animaux),
- une préparation de nerf dilacéré qui vous montrera des fibres nerveuses en long.
Le portail du Department of Biological Sciences - University of Delaware (Dr. Roger C. Wagner and Dr. Fred E. Hossler) vous fournira aussi d'excellentes vues microscopiques:
- coupe transversale de nerf périphérique traité à l'acide osmique (faible grossissement),
- coupe transversale de fibres myélinisées dans un nerf périphérique traité à l'acide osmique (fort grossissement),
- fibres dilacérées d'un nerf périphérique montrant les noeuds de Ranvier (fort grossissement),
Vous réaliserez des dessins de vos observation de manière à relater la structure d'un nerf et de ses fibres.
la moelle épinière lombaire, centre nerveux réflexe. Son organisation anatomique et notamment ses relations avec chacun des nerfs rachidiens pairs, auxquels elle est reliée par deux racines, l'une dorsale, comportant un ganglion rachidien et l'autre ventrale, peut être observée sur une coupe transversale observée à la loupe ou au très faible grossissement du microscope.
![]()
![]()
![]()
![]() Vous observerez au microscope une coupe transversale de moelle épinière à différents grossissements.
Vous observerez au microscope une coupe transversale de moelle épinière à différents grossissements.
Le portail du Département d'Histologie et d'embryologie de la faculté Universitaire FUNDP de NAMUR, dans lequel vous sélectionnerez la rubrique "Histologie générale - Tissus nerveux - Architecture schématique - La moelle épinière", vous permettra d'être guidé dans votre étude.
les muscles eux-mêmes, dont les myofibres avec leur plaque motrice individuelle, constituent les effecteurs. Il est ainsi possible de définir ce qu'est une unité motrice: c'est l'ensemble constitué par un neurone et ses ramifications, assurant la commande synchrone d'un certain nombre de fibres musculaires. Le microscope électronique permet de comprendre l'intimité de la jonction neuro-musculaire (synapse).
Tous ces éléments sont intégrés dans un circuit constituant l'arc réflexe, de type médullaire ici, dont nous allons justifier de l'intervention.
1.2 LES CIRCUITS NERVEUX DU RÉFLEXE
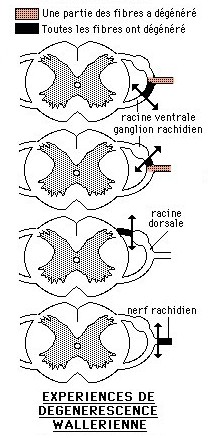 1.2.1 Les expériences de dégénérescence wallérienne:
1.2.1 Les expériences de dégénérescence wallérienne:
Ces expériences historiques ont été réalisées par Augustus Desiré WALLER en 1850.
Lorsque le corps cellulaire d'un neurone est détruit, la fibre nerveuse dégénère totalement. Lorsque la fibre nerveuse est sectionnée, la partie distale, c'est à dire la partie qui est séparée du corps cellulaire, dégénère. C'est le phénomène de la dégénérescence wallérienne. Par contre la partie proximale, c'est à dire celle qui est restée en rapport avec le corps cellulaire, peut régénérer.
![]() Waller a réalisé chez l'animal des sections de tronçons nerveux, notamment celles représentées sur les schémas ci-contre à droite. Donnez une interprétation aux résultats de chacune d'elles dans le but de préciser finalement où se trouvent les corps cellulaires des neurones intervenant dans le réflexe médullaire.
Waller a réalisé chez l'animal des sections de tronçons nerveux, notamment celles représentées sur les schémas ci-contre à droite. Donnez une interprétation aux résultats de chacune d'elles dans le but de préciser finalement où se trouvent les corps cellulaires des neurones intervenant dans le réflexe médullaire.
1.2.2 Les expériences de Bell-Magendie:
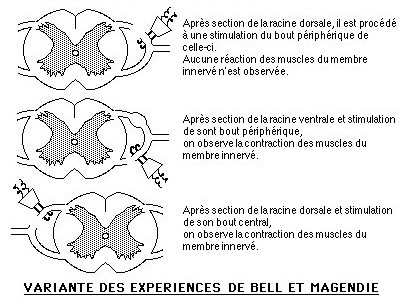 Charles BELL, dès 1811, entreprit de définir le rôle des racines rachidiennes dans la conduction nerveuse.
Charles BELL, dès 1811, entreprit de définir le rôle des racines rachidiennes dans la conduction nerveuse.
C'est François MAGENDIE en 1822 qui précisa le rôle des racines antérieures et postérieures des nerfs rachidiens, Charles BELL le confirma en 1824.
Une variante de ces expériences est présentée ci-contre à gauche: elles consistent à opérer, après anesthésie des seuls centres supérieurs (cerveau) chez un animal (animal spinal), des sections de racines rachidiennes, de les stimuler et d'observer les effets de ces stimulations.
![]() Analysez les résultats de ces expériences et tirez les conclusions qui s'imposent.
Analysez les résultats de ces expériences et tirez les conclusions qui s'imposent.
1.2.3 Deux types de neurones sont mobilisés:
Il est à présent possible d'identifier les deux types de neurones intervenant obligatoirement dans un réflexe:
- les neurones sensitifs afférents dont les influx centripètes cheminent des récepteurs musculaires à la moelle en passant par les racines dorsales des nerfs rachidiens: ils ont leurs péricaryons (corps cellulaires nucléés) dans les ganglions rachidiens.
![]()
![]()
![]()
![]() Là encore le portail du Département d'Histologie et d'embryologie de la faculté Universitaire FUNDP de NAMUR, dans lequel vous sélectionnerez la rubrique "Histologie générale - Tissus nerveux - Neurones - § Les cellules nerveuses ganglionnaires des ganglions rachidiens ou spinaux" vous permettra de reconnaître les neurones ganglionnaires en T.
Là encore le portail du Département d'Histologie et d'embryologie de la faculté Universitaire FUNDP de NAMUR, dans lequel vous sélectionnerez la rubrique "Histologie générale - Tissus nerveux - Neurones - § Les cellules nerveuses ganglionnaires des ganglions rachidiens ou spinaux" vous permettra de reconnaître les neurones ganglionnaires en T.
Vous en ferez un dessin d'observation, après avoir réalisé une observation sur une préparation du commerce.
- les motoneurones efférents dont les influx centrifuges empruntent les racines ventrales des nerfs rachidiens et parviennent aux muscles: leurs péricaryons se situent dans les cornes antérieures de la moelle (neurones multipolaires).
![]()
![]()
![]()
![]() Le même portail FUNDP et la même rubrique , mais § "Motoneurone" vous permettront de réaliser un dessin d'observation d'un neurone multipolaire.
Le même portail FUNDP et la même rubrique , mais § "Motoneurone" vous permettront de réaliser un dessin d'observation d'un neurone multipolaire.
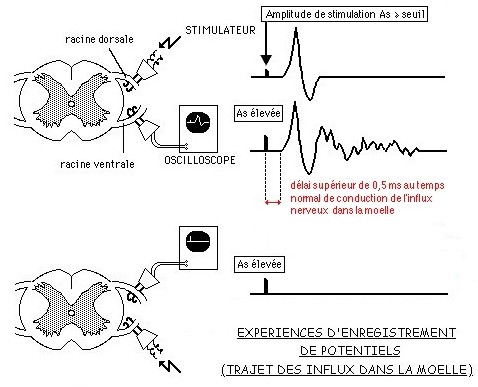 1.2.4 La connexion entre ces neurones se fait directement par des contacts synaptiques entre les axones des neurones en T et les dendrites ou les péricaryons des motoneurones: circuits monosynaptiques.
1.2.4 La connexion entre ces neurones se fait directement par des contacts synaptiques entre les axones des neurones en T et les dendrites ou les péricaryons des motoneurones: circuits monosynaptiques.
Il est possible de les mettre en évidence par des expériences d'enregistrements de potentiels.
Celles-ci consistent, après sections des racines rachidiennes et stimulation électrique, de rechercher le passage éventuel d'un message nerveux sur l'autre racine grâce à des électrodes réceptrices reliée à un oscilloscope ou un dispositif d'ExAO (voyez la figure ci-contre->).
![]() Analysez ces expériences et émettez des hypothèses sur les variantes des réponses obtenues, notamment en vous interrogeant sur les raisons du délai obtenu dans les deux premiers enregistrements.
Analysez ces expériences et émettez des hypothèses sur les variantes des réponses obtenues, notamment en vous interrogeant sur les raisons du délai obtenu dans les deux premiers enregistrements.
![]()
![]() Rendez compte par un schéma, en vous inspirant de ce document, des circuits nerveux plus complexes impliqués dans les expériences d'enregistrement de potentiel ci-dessus.
Rendez compte par un schéma, en vous inspirant de ce document, des circuits nerveux plus complexes impliqués dans les expériences d'enregistrement de potentiel ci-dessus.
1.3 ACTIVITE COORDONNEE DES MUSCLES ANTAGONISTES
1.3.1 La mobilité des articulations nécessite l'activité coordonnée de muscles antagonistes:
dans le réflexe achilléen, au muscle extenseur du pied (triceps sural) s'oppose un muscle fléchisseur (jambier antérieur): comment se comportent-ils lors de la mise en oeuvre du réflexe?
![]()

![]() Vous réaliserez l'ExAO suivante: trois électrodes sont placées sur le muscle du mollet (triceps sural = extenseur du pied) et trois autres sur le muscle situé à l'avant de la jambe (jambier antérieur = fléchisseur du pied) comme l'indique le dessin représenté par l'icône ci-contre à droite.
Vous réaliserez l'ExAO suivante: trois électrodes sont placées sur le muscle du mollet (triceps sural = extenseur du pied) et trois autres sur le muscle situé à l'avant de la jambe (jambier antérieur = fléchisseur du pied) comme l'indique le dessin représenté par l'icône ci-contre à droite.
Le réflexe est déclenché en frappant à l'aide du marteau qui est connecté au dispositif d'ExAO comme dans le § 1.1.1. Vous imprimerez l'enregistrement obtenu.
Observez le document ci-dessous:
Expliquez en quoi cet enregistrement montre que l'activité des muscles antagonistes est coordonnée.
1.3.2 Cette coordination est assurée par l'innervation réciproque de ces muscles antagonistes grâce à une double action des messages afférents, au niveau de la moelle:
- activation des motoneurones du muscle étiré, déclenchant une contraction,
- inhibition des motoneurones du muscle antagoniste, provoquant la chute de son tonus.
Le circuit excitateur monosynaptique est donc doublé d'un circuit inhibiteur polysynaptique (des interneurones ou neurones d'association s'intercalent entre neurones afférents et motoneurones).
![]()
![]() Complétez le schéma ci-contre à droite "coordination du réflexe rotulien" par des annotations.
Complétez le schéma ci-contre à droite "coordination du réflexe rotulien" par des annotations.
1.4 ROLE INTEGRATEUR DES MOTONEURONES MEDULLAIRES
Des messages provenant des centres nerveux supérieurs (cerveau) peuvent modifier l'amplitude de la réponse réflexe.
En effet une contraction volontaire du jambier antérieur (fléchisseur) inhibe le muscle antagoniste extenseur: l'étirement de celui-ci par percussion du tendon d'Achille se manifeste par une atténuation, voire une abolition du réflexe achilléen (courbe violacée sur l'enregistrement ci-dessous).
Les motoneurones sont donc capables d'intégrer deux informations contradictoires: l'excitation provenant des récepteurs musculaires étirés et l'inhibition d'origine cérébrale.
A l'inverse, si l'on utilise la manoeuvre de Jendrassik (méthode qui consiste à favoriser le réflexe en saisissant ses deux mains par les doigts fléchis et en les tirant de toutes ses forces) on constate une augmentation de l'amplitude de la réponse musculaire (courbe en vert ci-dessous).

Ces expériences témoignent du fait que les réflexes, s'ils font partie du patrimoine génétique de l'individu - ce que l'on nomme instinct chez l'animal - peuvent être modulés par l'intervention des centres encéphaliques.
Thème 2: Les messages nerveux au niveau neuronique
La communication nerveuse se manifeste par des signaux de nature électrique constituant des messages. L'activité électrique d'un nerf est facilement enregistrable en plaçant à son contact des électrodes reliées à un dispositif d'enregistrement. Celle des cellules nerveuses nécessite des techniques plus délicates.
2.1 LE SIGNAL ELEMENTAIRE DU MESSAGE NERVEUX, LE POTENTIEL D'ACTION
2.1.1 Approche expérimentale de l'activité neuronique: l'activité électrique du nerf de crabe
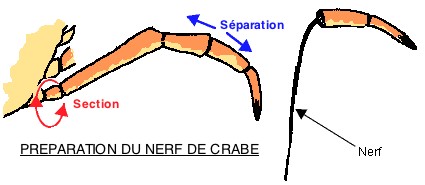
Principe:
Lorsqu'un nerf est en activité, il est parcouru par des messages nerveux se traduisant par une activité électrique globale qu'il est possible d'enregistrer après l'avoir amplifiée.
L'utilisation du crabe constitue une opportunité d'avoir un matériel approprié pour ce type de travaux en raison de la facilité à se procurer ces animaux à un faible coût et du peu de difficulté à isoler les nerfs sensorimoteurs des pattes.
Les réponse à divers types de stimulations électriques peuvent être visualisées sur un écran d'oscilloscope ou avec un système d'ExAO. Il est ainsi possible de déterminer:
- le seuil d'excitabilité du nerf et de ses différents groupes de fibres,
- leur vitesse de conduction.
Les portails BIOMEDIA de l'université de Jussieu ou ceux de SVT de l'Académie de Lille, ou de Rennes donnent un aperçu de la façon de procéder, notamment pour extirper le nerf.
Protocole et montage:
Sur un crabe, on fait plier l'avant dernière articulation d'une patte de manière à briser la carapace. On étire les deux articles distaux dans l'axe de la patte: le nerf y reste accroché.
Une fois le nerf disséqué et placé sur les électrodes d'une table à nerf alimentée par du liquide physiologique ou de l'eau de mer reconstituée, on relie 2 électrodes de stimulation et un fil de masse à l'interface (face avant sur le document ci-contre) et deux électrodes de réception à l'interface (latéralement sur le même document).
Enregistrements:

![]()

![]() Le matériel peut être testé à l'aide d'un fil de coton imbibé de liquide physiologique. Une stimulation peut être effectuée, un artefact se voit alors, ce qui montre que celui-ci est lié au courant de stimulation et non à la réponse du nerf.
Le matériel peut être testé à l'aide d'un fil de coton imbibé de liquide physiologique. Une stimulation peut être effectuée, un artefact se voit alors, ce qui montre que celui-ci est lié au courant de stimulation et non à la réponse du nerf.
1 - On étudie dans un premier temps la réponse du nerf à une stimulation efficace: pour ce faire on stimule le nerf avec une forte amplitude, en choisissant pour commencer des électrodes réceptrices et stimulatrices rapprochées (5 mm).
Observons un tel enregistrement. Définissez les caractéristiques du potentiel global: amplitude en mV, durée en ms.
Calculez la vitesse de propagation du message nerveux.
2 - Faites varier l'amplitude de la stimulation et superposez les enregistrements obtenus. Regardez comment varie l'amplitude de la réponse globale du nerf.
En utilisant les observations histologiques faites sur la structure d'un nerf, proposez une explication aux variations constatées. Pouvez-vous définir ce que l'on appelle seuil d'excitabilité du nerf?
2.1.2 Exploration électrique d'une fibre isolée:
La membrane au repos est polarisée électriquement:
La nature et l'origine de l'activité des nerfs est à rechercher au niveau des fibres nerveuses.
 L'expérience que nous allons simuler est par exemple réalisée sur une fibre nerveuse géante (axone de calmar dont le diamètre peut atteindre 1mm).
L'expérience que nous allons simuler est par exemple réalisée sur une fibre nerveuse géante (axone de calmar dont le diamètre peut atteindre 1mm).
Lorsqu'une fibre émet, conduit et transmet un message nerveux, elle est dite "en activité", une fibre vivante qui n'est pas en activité est dite "au repos". Celle qui se trouve dans la boîte à électrodes restera au repos tout au long de cette expérience: noter en effet l'absence d'électrodes de stimulation destinées à la mettre en activité.
Les électrodes branchées à l'oscilloscope sont des électrodes de détection: l'une est une électrode de référence (à la masse) ou placée à la surface externe de la membrane plasmique de la fibre (électrode de surface), l'autre sera introduite à l'intérieur du cytoplasme (microélectrode).
![]()
![]() Cliquez sur l'icône ci-contre, liée à l'animation ->
Cliquez sur l'icône ci-contre, liée à l'animation ->
Phase 1: La microélectrode n'est pas en contact avec la fibre, elle est à l'extérieur de la membrane plasmique. Sur l'écran de l'oscilloscope la ligne horizontale graduée correspond à la valeur O du potentiel, chaque grande division verticale vaut 20 mV.
Phase 2: En cliquant sur le bouton ![]() vous enfoncerez la microélectrode au travers de la membrane et l'oscilloscope enregistrera alors la variation de potentiel entre les deux électrodes.
vous enfoncerez la microélectrode au travers de la membrane et l'oscilloscope enregistrera alors la variation de potentiel entre les deux électrodes.
Chiffrez et interprétez la variation de potentiel (notez bien le branchement des deux électrodes avec les plaques horizontales de l'oscilloscope: ce sont elles qui enregistrent toute variation de potentiel entre les deux électrodes, par déviation du faisceau d'électrons).
Ces expériences ne sont pas réalisables dans un environnement de lycée, car elles demandent d'utiliser un matériel très perfectionné, mais il est possible, en classe, de mesurer le potentiel de repos d'une cellule-œuf, celui de la poule, en "bricolant" un dispositif d'enregistrement.
Le potentiel d'action, un événement membranaire brutal:L'expérience est réalisée sur le même matériel que précédemment et avec le même dispositif, l'électrode de surface n'a pas bougé, la microélectrode reste implantée à l'intérieur de la fibre: l'expérience est la suite chronologique de la précédente (enregistrement du potentiel de repos). Mais cette fois-ci la fibre va être mise en activité grâce à deux électrodes de stimulation S1 et S2 (S1 est l'anode, S2 la cathode) reliées à un dispositif de stimulation.
![]()
![]() La fréquence d'excitation que l'on choisira sera de 25 hz, ce qui permet, grâce au dispositif de synchronisation et à la persistance des impressions lumineuses, de voir le phénomène en permanence. Jusqu'à 100 hz la fréquence n'a pas d'influence notable sur la réponse nerveuse. Au delà la période réfractaire intervient.
La fréquence d'excitation que l'on choisira sera de 25 hz, ce qui permet, grâce au dispositif de synchronisation et à la persistance des impressions lumineuses, de voir le phénomène en permanence. Jusqu'à 100 hz la fréquence n'a pas d'influence notable sur la réponse nerveuse. Au delà la période réfractaire intervient.
Le bouton Fs reste donc fixe au cours de l'expérience (en blanc sur le schéma).
On choisit une valeur fixe de la durée de stimulation ts (bouton en blanc sur le schéma).
L'expérience consiste à stimuler la fibre en envoyant des stimulations d'amplitude croissante (bouton As en noir sur le schéma): on enregistre les potentiels d'action à l'écran de l'oscilloscope.
Les conditions de la naissance d'un potentiel d'action
A l'aide du dispositif ci-dessus on soumet une fibre à plusieurs stimulations électriques de durée constante et d'amplitude régulièrement croissante. On observe sur l'écran de l'oscilloscope les réponses correspondantes de la fibre.
![]()
![]() Vous pouvez réaliser cette simulation en utilisant le même logiciel "PREPOS" que dans l'expérience précédente.
Vous pouvez réaliser cette simulation en utilisant le même logiciel "PREPOS" que dans l'expérience précédente.
En neurophysiologie on dit que la fibre nerveuse obéit à la "loi du tout ou rien": le potentiel d'action qui naît et se propage présente d'emblée son amplitude maximale dès que la fibre est dépolarisée pour et au-delà d'un certain "seuil de stimulation".
En observant les enregistrements obtenus, précisez ce que l'on entend par "seuil de dépolarisation" et donnez-en une valeur approximative. Lorsque ce seuil est dépassé, observez-vous une variation de l'amplitude de la réponse nerveuse? Quelle caractéristique du signal nerveux pouvez-vous ainsi évoquer?

Les activités du programme relatées dans les deux § suivants nécessitent de faire appel à des expérimentations irréalisables dans un établissement scolaire. Un logiciel de simulation "Récepteur, neurone, synapse" mis au point par François TILQUIN en 2000 - Académie de GRENOBLE - remédie à cet état de fait.
Vous pouvez le télécharger sur le site http://www.ac-grenoble.fr/svt/old_site/logiciel/neuro/neuro.htm
2.2 NAISSANCE, CODAGE ET CONDUCTION DES MESSAGES NERVEUX:
2.2.1 Un exemple de message: le message sensoriel:
Dans l'organisme, les signaux nerveux prennent naissance au niveau de structures spécifiques, les récepteurs sensoriels. Le fuseau neuro-musculaire en est un exemple, la peau en contient plusieurs types.
Les récepteurs ont une sensibilité en réagissant à un stimulus précis.
Ce sont des capteurs pouvant détecter des paramètres provenant soit du milieu extérieur, soit de l'intérieur de l'organisme.
Suivant le type de récepteur, le stimulus est d'ordre mécanique, thermique, chimique, électrique, lumineux, et déclenche la stimulation de neurones sensitifs émettant des messages en direction d'un centre nerveux.
Ils présentent également une certaine excitabilité: il existe un seuil d'excitation en deçà duquel un récepteur n'émet aucun potentiel d'action (stimulus infra-liminaire), mais seulement une faible dépolarisation locale qui ne se propage pas, le potentiel de récepteur.
Un potentiel d'action ne se déclenche que pour une valeur minimale de la stimulation ou seuil et pour une valeur au delà du seuil (stimulation liminaire ou supra-liminaire).
Le message nerveux afférent est codé comme nous allons le constater.
2.2.2 Le double codage des messages nerveux:
![]()
![]() Le module "SYNAPSE" du logiciel présente plusieurs fibres de neurones afférents (n1 à n5) qui s'articulent par des synapses à un neurone efférent (n6). Des boutons (s1, ...) permettent de porter des stimulations avec des intensités variables.
Le module "SYNAPSE" du logiciel présente plusieurs fibres de neurones afférents (n1 à n5) qui s'articulent par des synapses à un neurone efférent (n6). Des boutons (s1, ...) permettent de porter des stimulations avec des intensités variables.
Différentes activités sont proposées.
Afin de mettre en évidence la manière dont se fait le codage de l'information nerveuse, vous pouvez porter sur un ensemble de fibres nerveuses (n3, n4, n5), pouvant représenter un nerf sensoriel, une série de stimuli d'intensité croissante et vous observerez comment varie la réponse sur le neurone n6.
Dans tous les cas, les messages nerveux sont des salves ou trains de potentiels d'action, dont l'amplitude est constante, mais dont la fréquence est variable.
Au niveau d'une fibre le message est donc codé en "modulation de fréquence": généralement la fréquence augmente quand l'intensité du stimulus augmente.
![]()
![]() Calculez les fréquences des réponses obtenues ci-dessus (en herz = nombre de potentiels d'action par seconde).
Calculez les fréquences des réponses obtenues ci-dessus (en herz = nombre de potentiels d'action par seconde).
Au niveau d'un nerf, un deuxième codage, ayant une signification pour l'organe qui le reçoit, correspond au nombre de fibres mobilisées à un instant donné. C'est ce deuxième codage qui explique les variations d'amplitude du potentiel global d'un nerf.
Enfin, la durée d'un message représente aussi une information pertinente: elle peut informer un centre de la durée d'un stimulus.
2.2.3 La conduction des messages le long des fibres nerveuses: la conductibilité
Comme il a été précisé ci-dessus, vous pouvez donner libre cours à diverses activités à l'aide du logiciel pour illustrer les propriétés du tissu nerveux.
Un potentiel d'action n'affecte à un instant donné qu'une zone limitée de la membrane nerveuse,
Il se propage rapidement,
Son amplitude reste constante d'une extrémité à l'autre de la fibre.
La vitesse de propagation est variable d'une fibre à l'autre: de 1 à plus de 100m.s-1.
Celle-ci dépend du calibre et de la nature des fibres: elle est d'autant plus grande que la fibre est plus grosse et les fibres à myéline sont plus rapides que celles qui en sont dépourvues.
2.3 LA TRANSMISSION SYNAPTIQUE ET L’INTÉGRATION DES MESSAGES NERVEUX
Le message nerveux, en arrivant à l'extrémité de l'axone, atteint une zone de connexion avec une autre cellule (neurone, myofibre,...), appelée synapse, caractérisée par:
une transmission unidirectionnelle (cellule pré- et cellule post-synaptique),
un délai synaptique de l'ordre de 0,5 ms (cf. §1.2.4).
Trois animations Flash à propos de la synapse sont disponibles sur le site: http://www.ac-creteil.fr/svt/acc_lyc.htm à la rubrique "1èreS - Médias - Consulter".
Vous pouvez aussi les télécharger en cliquant directement sur l'icône ci-contre à droite.
Trois activités sont proposées:
![]() Ultrastructure
Ultrastructure ![]() Expérimentation
Expérimentation ![]() Fonctionnement
Fonctionnement
2.3.1 Transmission chimique des messages nerveux:
La cellule pré-synaptique est séparée de la cellule post-synaptique par un espace ou fente synaptique de 20 à 50 nm. Son franchissement ne peut pas se faire directement, mais est assuré par une étape chimique. De nombreuses vésicules remplies d'un neuro-transmetteur ou médiateur chimique se trouvent dans le cytoplasme de la terminaison axonique.
![]()
![]()
![]() En utilisant l'option "Ultrastructure" vous accéderez à une électronographie qui vous révélera la configuration d'une synapse neuro-neuronique. Faites un dessin d'observation de celle-ci.
En utilisant l'option "Ultrastructure" vous accéderez à une électronographie qui vous révélera la configuration d'une synapse neuro-neuronique. Faites un dessin d'observation de celle-ci.
L'arrivée de potentiels d'action y déclenche la libération par exocytose d'un nombre plus ou moins important de molécules de neuro-transmetteur. Celui-ci se lie à des récepteurs spécifiques de la membrane post-synaptique, déclenchant ainsi la naissance d'un nouveau message. La destruction enzymatique rapide du neuro-transmetteur et la recapture des molécules interrompt la transmission.
![]()
![]() Visualisez les animations "Expérimentation" et "Fonctionnement" et relevez la chronologie des événements intervenant dans la transmission synaptique.
Visualisez les animations "Expérimentation" et "Fonctionnement" et relevez la chronologie des événements intervenant dans la transmission synaptique.
2.3.2 Synapses excitatrices et synapses inhibitrices:
le principe de fonctionnement des synapses est identique, mais selon le neuro-transmetteur qu'elle libère et qui est toujours le même pour une synapse donnée, l'effet sur le neurone post-synaptique peut-être:
excitateur: naissance d'un nouveau message,
inhibiteur: annulation ou diminution de l'émission de potentiels d'action.
![]()
![]()
![]() Parmi les neurones afférents du module "SYNAPSE", qui s'articulent avec le neurone efférent n6, tous n'ont pas le même effet. Sollicitez-les et voyez comment se présente le message transmis. Imprimez ou schématisez les réponses obtenues et précisez les effets sur la genèse des potentiels d'action véhiculés par le neurone efférent.
Parmi les neurones afférents du module "SYNAPSE", qui s'articulent avec le neurone efférent n6, tous n'ont pas le même effet. Sollicitez-les et voyez comment se présente le message transmis. Imprimez ou schématisez les réponses obtenues et précisez les effets sur la genèse des potentiels d'action véhiculés par le neurone efférent.
2.3.3 Intégration des messages:
Dans un centre nerveux, un neurone peut recevoir des milliers de terminaisons axoniques par l'intermédiaire de son péricaryon ou ses dendrites.
![]()
![]()
![]() Vous procéderez aux expérimentations suivantes:
Vous procéderez aux expérimentations suivantes:
stimulation répétée d'une fibre afférente (sommation temporelle),
stimulation de plusieurs fibres afférentes (sommation spatiale),
et vous observerez à chaque fois les effets de celles-ci sur le neurone efférent post-synaptique.
Soit excitatrices, soit inhibitrices, le neurone efférent intègre toutes les informations, même contradictoires, pour en faire la "somme algébrique": si le résultat est une dépolarisation suffisante de la membrane post-synaptique, des potentiels d'action sont émis par le neurone post-synaptique, sinon il reste au repos.
Cette fonction intégratrice joue un rôle essentiel dans le traitement des messages transitant dans un centre nerveux.
2.3.4 Application au réflexe myotatique achilléen:
Suite à l'étirement du muscle extenseur (soléaire), l'activité des motoneurones du muscle extenseur est augmentée sous l'influence de synapses excitatrices, celle des motoneurones du muscle fléchisseur est diminuée sous l'influence de synapses inhibitrices.
Mais il est possible de concevoir, lors de l'atténuation ou de l'abolition du réflexe par une contraction volontaire du muscle fléchisseur, que les motoneurones du muscle extenseur reçoivent des informations inhibitrices par l'intermédiaire d'interneurones.
![]() Revoyez à ce sujet le schéma des circuits nerveux du §1.3.2 pour bien comprendre cette notion d'intégration des messages.
Revoyez à ce sujet le schéma des circuits nerveux du §1.3.2 pour bien comprendre cette notion d'intégration des messages.
Vous pouvez réviser les notions abordées dans le thème 2 en utilisant le logiciel "Nerf" téléchargeable à l'adresse suivante: http://pedagogie.ac-toulouse.fr/svt/serveur/lycee/perez/nerf/nerf.htm
Thème 3: la plasticité du système nerveux central
 La dissection de la souris révèle l'organisation du système nerveux d'un mammifère.
La dissection de la souris révèle l'organisation du système nerveux d'un mammifère.
L'encéphale de l'homme, contenu dans la boîte crânienne, est constitué de l'avant vers l'arrière et du haut en bas:
des hémisphères cérébraux (appelés communément "cerveau") dont le cortex ou écorce forme la partie superficielle,
du cervelet,
du tronc cérébral dont le bulbe rachidien.
![]() Entraînez-vous à reconnaître les différentes parties de l'encéphale sur les photographies ci-contre à gauche.
Entraînez-vous à reconnaître les différentes parties de l'encéphale sur les photographies ci-contre à gauche.
Nous aborderons la fonction de réception et de traitement des informations sensorielles du cortex cérébral.
3.1 LA RÉCEPTION DES MESSAGES SENSORIELS PAR LE CORTEX CÉRÉBRAL:
3.1.1 Grâce à l'imagerie médicale (idéographie, tomographie densitométrique, imagerie par résonance magnétique IRM), nous pouvons identifier les régions du cortex.
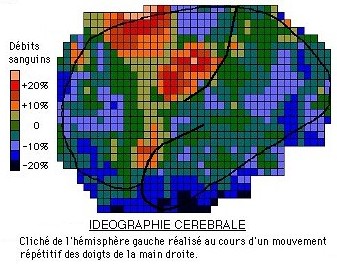
L'idéographie est une technique d'exploration du fonctionnement cérébral et des processus mentaux par mesure des débits sanguins locaux à la surface du cortex cérébral: ils permettent de localiser les zones actives lorsqu'un sujet effectue une fonction motrice ou sensorielle (vue, ouïe, toucher,...) et ainsi de dresser la cartographie des aires d'arrivée des messages sensoriels ou de départ des signaux moteurs.
Il est possible de cartographier des aires corticales motrices (en rouge) et sensorielles (en bleu) comme le montre le document ci-dessous.
 3.1.2 Exemple de l'aire somato-sensorielle qui traite les informations tactiles.
3.1.2 Exemple de l'aire somato-sensorielle qui traite les informations tactiles.
Comme la plupart des aires, elle se subdivise en:
une aire primaire de projection située dans la circonvolution à l'arrière du sillon de Rolando: elle correspond au point d'arrivée des messages provenant des différentes parties du corps,
une aire secondaire d'association, en arrière de la précédente: elle assure la connexion de l'aire de projection avec de nombreuses autres régions du cerveau.
![]() Localisez sur l'idéogramme ci-dessus l'activité cérébrale la plus intense lors d'un mouvement répétitif des doigts de la main droite. Quelle remarque pouvez-vous faire quant au trajet des voies nerveuses issues de l'encéphale?
Localisez sur l'idéogramme ci-dessus l'activité cérébrale la plus intense lors d'un mouvement répétitif des doigts de la main droite. Quelle remarque pouvez-vous faire quant au trajet des voies nerveuses issues de l'encéphale?
Chaque hémisphère reçoit l'ensemble des informations sensorielles de la moitié opposée du corps sur une représentation appelée " homunculus sensoriel ", dont l'étendue des différentes parties est liée à la densité des terminaisons nerveuses correspondantes. Cette projection corticale varie d'une espèce à l'autre en fonction de la sensibilité de telle ou telle partie du corps:
chez l'homme, la main est bien représentée,
chez le singe, arboricole, le pied est presque aussi bien représenté que la main,
chez le rat, nocturne, les vibrisses sont les organes des sens les mieux représentés.
Le schéma ci-contre explique comment il faut concevoir cette représentation corticale dans le cas de la sensibilité tactile chez l'homme.
3.2 ORGANISATION DES NEURONES CORTICAUX
3.2.1 Six couches de neurones:
Le cortex cérébral renferme dans son épaisseur de 2 à 4 mm environ 100 milliards de neurones.
Par son axone, chaque neurone communique avec un nombre limité d'autres neurones, mais le péricaryon et les dendrites peuvent recevoir plusieurs milliers de connexions.
L'observation microscopique révèle six couches superposées (numérotées de I à VI vers l'intérieur). Les neurones pyramidaux y dominent: leur structure révèle une arborisation dendritique et un axone.
![]()
![]() Vous pouvez mener une étude histologique du cortex cérébral sur le lien suivant:
Vous pouvez mener une étude histologique du cortex cérébral sur le lien suivant:
http://brainmaps.org/index.php?q=cerebral%20cortex et notamment avec la préparation n°8 ![]()
Les 6 couches de I à VI se caractérisent ainsi de la surface vers la profondeur :
- la couche I ou couche moléculaire pauvre en cellules nerveuses, constituée surtout des cellules gliales et des fibres nerveuses à trajets parallèles à la surface corticale.
- la couche II ou couche granulaire externe, essentiellement constituée de petits neurones non pyramidaux.
Remarque : chez les rongeurs, ces deux couches (I et II) n'en forment qu'une.
- La couche III ou couche pyramidale externe, contient la majorité des neurones pyramidaux. Les axones de ces neurones forment les fibres intrahémisphériques et interhémisphériques ou commissurales dans la substance blanche tandis que les dendrites apicales atteignent la couche moléculaire.
- La couche IV ou couche granulaire interne formée de petits neurones non pyramidaux représentés essentiellement par des cellules étoilées.
- La couche V ou couche pyramidale interne contient essentiellement des neurones pyramidaux dont les dendrites apicales se projettent soit dans la couche moléculaire (couche I) soit dans la couche granulaire interne (couche IV). Les axones de ces neurones se projettent à distance (striatum, tronc cérébral, moelle épinière..) contrairement à ceux de la couche III.
- La couche VI ou multiforme est la source principale des neurones qui se projettent sur le thalamus. Elle est à l'origine aussi des fibres commissurales et des fibres d'association.
3.2.2 Une organisation "en colonnes" du cortex sensoriel:
En descendant une microélectrode perpendiculairement à la surface du cortex il est possible de montrer que toutes les cellules rencontrées répondent à des stimulations mécaniques de même type, appliquées au même endroit de la surface du corps, mais pas pour des stimulations appliquées dans des régions voisines (travaux de MOUNCASTLE dans les années 1960).
Si la microélectrode est introduite tangentiellement, les neurones atteints répondent pour des stimuli appliqués en des points différents de la peau.
Par ailleurs, le marquage des fibres afférentes, apportant des informations sensorielles, montre qu'elles aboutissent toutes dans la couche IV, qui constitue la "porte d'entrée" du cortex.
Le cortex est ainsi subdivisé en modules élémentaires constitué en colonnes ou "tonneaux" ("barrels" en anglais) de 2 à 3 mm d'épaisseur et d'environ 300 µm de diamètre, dont l'entrée des informations (en provenance des noyaux relais thalamiques spécifiques) se fait par les neurones de la couche IV et la sortie par les neurones des autres couches vers les autres régions corticales et sous-corticales.
Il existe un certain degré de traitement de l'information au sein de chaque colonne corticale. Certains neurones du cortex primaire sont capables de détecter la direction du stimulus dans le champ récepteur. Ils détectent également l'intensité du stimulus: l'intensité de la décharge neuronale est fonction de l'intensité de la stimulation du champ récepteur périphérique. La colonne est l'unité fonctionnelle de base du cortex cérébral.
Le cortex sensitif est donc constitué par la juxtaposition d'innombrables colonnes corticales qui constitue la carte somatotopique du corps (" homonculus sensitif").
Cette représentation corticale peut particulièrement être étudiée sur un territoire corporel contenant de nombreux récepteurs sensoriels comme par exemple les vibrisses des rongeurs.
Peu d'études mettent actuellement en évidence la réorganisation permanente de l'architecture neuronale dans le cerveau d'animaux adultes au cours d'un comportement naturel.
![]()
![]() Vous pouvez découvrir à ce sujet l'étude réalisée sur les changements répétés de la morphologie dendritique dans l'hippocampe de l'écureuil sibérien au cours de l'hibernation en cliquant sur l'icône à droite.
Vous pouvez découvrir à ce sujet l'étude réalisée sur les changements répétés de la morphologie dendritique dans l'hippocampe de l'écureuil sibérien au cours de l'hibernation en cliquant sur l'icône à droite.
3.3 LA PART DU GÉNOTYPE DANS LE FONCTIONNEMENT DU SYSTÈME NERVEUX
3.3.1 Dans l'exemple du réflexe myotatique qui est une réaction nerveuse involontaire, automatique et stéréotypée à l'étirement d'un muscle, la chaîne de neurones étant toujours organisée de la même façon, il faut admettre qu'elle est mise en place au cours du développement embryonnaire, sous le contrôle de l'information génétique.
Pour plus de détails sur la neurogénèse (développement du tissu nerveux) et plus précisément celle des cortex cérébral et cérébelleux, vous pouvez consulter la page:
http://www.inrp.fr/Acces/biotic/gpe/dossiers/mutcer/html/neurogenese.htm
3.3.2 Des preuves déduites de l'étude de l'effet de mutations chez des souris présentant des troubles moteurs (souris weaver, reeler,...) et chez l'homme, dans le cas de maladies rares (insensibilité congénitale à la douleur) ont pu être mises en évidence.
La mise en place, au cours du développement embryonnaire, de chaînes neuroniques dans le système nerveux central, comme dans le câblage périphérique est, au moins en partie, sous le contôle génétique.
3.4 UN REMODELAGE POSSIBLE DES CIRCUITS NERVEUX CHEZ L'ADULTE
3.4.1 La malléabilité de l'aire somato-sensorielle a pu être montrée chez l'animal comme chez l'homme (amputations, blessures): une réaffectation de l'aire de projection d'un organe au profit des aires de projection adjacentes est possible.
![]()
![]() Vous pouvez observer les résultats du remodelage cortical chez un patient atteint de syndactylie (soudure congénitale des doigts) qui a subi une intervention chirurgicale permettant la séparation des doigts.
Vous pouvez observer les résultats du remodelage cortical chez un patient atteint de syndactylie (soudure congénitale des doigts) qui a subi une intervention chirurgicale permettant la séparation des doigts.
Ces résultats sont mis en évidence par la technique de magnéto-encéphalographie (MEG) représentée par l'icône ci-contre. C'est une méthode de mesure et de cartographie des champs magnétiques extrêmement faibles produits par les neurones en action.
Vous pouvez également consulter un article sur la réorganisation cérébrale des régions motrices après une greffe des deux mains.
L'entraînement peut aussi avoir pour conséquence une extension de la projection d'un organe au détriment des aires de projection adjacentes (pouce et auriculaire des violonistes par exemple).
3.4.2 Les mécanismes de remodelage épigénétique qui tiennent à une modification de la quantité et de l'intensité des connexions synaptiques entre les neurones d'une colonne et ceux des colonnes adjacentes témoignent d'une plasticité remarquable du cortex, mais sont encore mal connus.