Morgan: théorie chromosomique de l'hérédité
MORGAN ET LA THEORIE CHROMOSOMIQUE DE L'HEREDITE
Publiés en 1866 dans un article des Comptes rendus de la Société d'histoire naturelle de Brno, les travaux de Mendel passent à peu près inaperçus, n'étant cités qu'une douzaine de fois entre leur publication et leur redécouverte en 1900. Cette redécouverte est due à Hugo de Vries (Amsterdam), Carl Correns (Berlin), et Erich Tschermack (Vienne), qui retrouvent de façon indépendante des résultats similaires à ceux de Mendel.
Le fait que ces travaux soient restés si longtemps sans impact s'explique sans doute en partie par plusieurs lacunes qu'ils laissent transparaître pour les contemporains. Tout d'abord, les travaux de Mendel ne permettent pas d'expliquer la réapparition d'un caractère qui existait non pas chez les parents ou grands-parents, mais chez un ancêtre qui peut être beaucoup plus éloigné. D'autre part, ces travaux ne concernent que les caractères à versions alternatives tranchées, et laissent complètement de côté les caractères à variations continues.
Par ailleurs, la découverte des chromosomes et des mécanismes de leur transmission va conduire à postuler que ceux-ci sont le support des unités d'information héréditaires - les "facteurs" mendéliens - que l'on appellera gènes.
Thèmes traités et activités pédagogiques (durée: 2 semaines):
![]() La redécouverte des lois de Mendel
La redécouverte des lois de Mendel
![]() La théorie chromosomique de l'hérédité
La théorie chromosomique de l'hérédité
![]() Les premières cartes génétiques et la définition provisoire du gène
Les premières cartes génétiques et la définition provisoire du gène
1 La redécouverte des lois de Mendel
1.1 LES "PANGÈNES" DE HUGO DE VRIES
En 1889, Hugo de VRIES est professeur de Botanique à Amsterdam. Il a à son actif plusieurs contributions importantes: l'une d'elles concerne une théorie de l'hérédité impliquant des particules élémentaires qu'il baptise "pangènes". Il se propose d'en étudier la transmission en réalisant des hybridations chez les végétaux entre 1890 et 1899.
Il publie en 1900, dans les Comptes rendus de l'Académie des Sciences de Paris, une note "Sur la loi de disjonction des hybrides".
Ayant réalisé un grand nombre de croisements chez des végétaux, il obtient, comme Mendel, à partir de lignées parentales pures (D = caractère dominant, r = caractère récessif), les résultats suivants:
|
P |
D |
r |
|
F1 |
Dr (100%) |
|
F2 |
D (25%) |
Dr (50%) |
r (25%) |
"Dans l'hybride, le caractère simple différentiel des parents est donc visible ou dominant, tandis que le caractère antagoniste est récessif. Ces caractères antagonistes restent ordinairement combinés pendant toute la vie végétative, [...] mais chaque grain de pollen et chaque ovule ne reçoit que l'un des deux.
[...] on peut représenter le nombre et la nature des hybrides par la formule représentative suivante, dans laquelle les nombres D et r sont égaux:
(D + r) (D + r) = De2 + 2Dr + re2
soit, dans le cas ci-dessus: (0,5 + 0,5) (0,5 + 0,5) = (0,25 + 0, 5 + 0,25):
|
0,5 D |
0,5 r | ||
|
0,5 D |
0,25 D |
0,25 Dr | ||
|
0,5 r |
0,25 Dr |
0,25 r |
[...] Les individus D auront le caractère dominant pur, l'ayant hérité du père et de la mère. De la même façon, les individus r auront le caractère récessif pur, tandis que Dr seront les hybrides. Ceux-ci porteront le caractère dominant apparent et le caractère récessif latent.
[...] L'ensemble de ces expériences met donc en évidence la loi de disjonction des hybrides et vient confirmer les principes que j'ai énoncés sur les caractères spécifiques considérés comme des unités distinctes."
1.2 LES LOIS DE MENDEL S'APPLIQUENT AUX ANIMAUX
Les travaux du français Cuenot sur la souris: Lucien CUENOT, professeur de zoologie à Nancy, reprend les lois de Mendel dont il est longtemps le seul en France à proclamer la valeur et montre l'application de ces lois à l'ensemble des espèces animales. En 1903-1906, il prouve que des caractères de coloration de la souris correspondent à divers états d'un même gène; ces états nés par mutation sont des alternatives multiples.
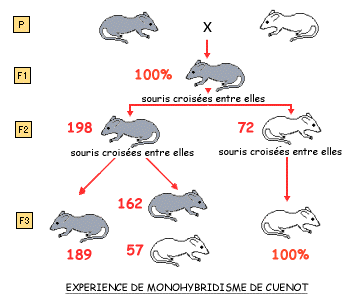
Il est possible de tester la nature des souris grises de F2 en réalisant des croisements-tests: ceux-ci consistent à croiser un individu dont on n'est pas sûr de la nature (souris grise hybride ou de lignée pure) avec un individu dont on est sûr de la nature génétique (souris blanche de lignée pure). Les résultats obtenus sont différents selon l'alternative envisagée.

![]()
![]() Dans les deux cas ci-dessus expliquez les résultats obtenus en dressant l'échiquier si nécessaire.
Dans les deux cas ci-dessus expliquez les résultats obtenus en dressant l'échiquier si nécessaire.
1.3 LES "LOIS DE MENDEL" DEFINIES PAR L'ALLEMAND CORRENS
Carl CORRENS rend hommage à MENDEL en utilisant l'expression "lois de MENDEL":
Première loi de Mendel ou loi de pureté des gamètes: au moment de la formation des gamètes, il y a ségrégation (= séparation) des unités distinctes qui portent les caractères différentiels, qui se réassocient au moment de la fécondation.
Seconde loi de Mendel ou loi de ségrégation indépendante des caractères: les deux unités d'un même couple de caractères ségrègent indépendamment de celles des autres couples.
2 La théorie chromosomique de l'hérédité
A la fin du XIXe siècle, les progrès réalisés en microscopie permettent de préciser les connaissances dans le domaine de la reproduction sexuée:
En 1875, HERTWIG observe la fécondation chez l'oursin (pénétration du spermatozoïde dans l'ovule et fusion des noyaux mâle et femelle),
Vers 1880 des "bâtonnets" que l'on nommera chromosomes sont identifiés dans le noyau,
En 1883 Van BENEDEN observe quatre chromosomes dans l'oeuf d'un Ascaris et seulement deux chromosomes dans les gamètes mâles et femelles.
2.1 LES ETUDES CYTOLOGIQUES DE SUTTON: LES CHROMOSOMES
Au début du XXe siècle des cytologistes découvrent la méiose dans les cellules de la lignée germinale des insectes, c'est-à-dire les cellules qui conduisent à la formation des gamètes (par opposition aux cellules somatiques, c'est-à-dire toutes les autres).
Dans un article publié en 1902 dans le laboratoire de zoologie de la Columbia University et intitulé "On the morphology of the chromosome group in Brachystola magna" Walter S. SUTTON rapporte ses études cytologiques sur les chromosomes de sauterelle qui l'amènent à conclure que, lors de la gamétogénèse:
les chromosomes ont une individualité,
qu'ils apparaissent sous forme de paires, avec un membre de chaque paire constitué par chaque parent,
que les chromosomes appariés se séparent l'un de l'autre pendant la méiose.
Il conclut ainsi: "je peux finalement attirer l'attention sur la probabilité que l'association des chromosomes paternels et maternels dans les paires et leur séparation pendant la réduction chromatique [...] peut constituer la base physique des lois de l'hérédité mendélienne."
Ils met en effet en évidence les chromosomes spiralisés, qu'il nomme "spirèmes": ceux-ci se groupent par paires au début de la méiose, qu'il appelle "bivalents". Chaque bivalent est formé par l'association d'un chromosome d'origine paternelle et d'un chromosome d'origine maternelle.
L'observation du déroulement de la méiose lui permet de constater que le chromosome paternel et le chromosome maternel de chaque bivalent se séparent au hasard à chaque pôle de la cellule de manière à ce que les gamètes héritent chacun d'une combinaison de chromosomes paternels et maternels et celle-ci n'est pas la même selon le gamète considéré.
Cette déduction logique de la part de SUTTON ne peut être vérifiée à l'époque, car il avoue ne pas pouvoir reconnaître le chromosome paternel du chromosome maternel.
![]() Faites le lien entre les observations de SUTTON et les lois de MENDEL. Précisez pourquoi les propositions de SUTTON ne constituaient, à l'époque, qu'une théorie.
Faites le lien entre les observations de SUTTON et les lois de MENDEL. Précisez pourquoi les propositions de SUTTON ne constituaient, à l'époque, qu'une théorie.
En partant des échiquiers de croisement F1xF1 obtenus par MENDEL dans le monohybridisme et le dihybridisme, déterminez les combinaisons chromosomiques qui résultent de la méiose et de la fécondation dans le tableau ci-dessous:
|
Nombre diploïde de chromosomes |
Nombre haploïde de chromosomes |
Combinaisons gamétiques différentes |
Combinaisons zygotiques (oeuf) différentes |
|
2 |
|
|
|
|
4 |
|
|
|
|
6 |
|
|
|
|
n |
|
|
|
2.2 LA FORMULATION DE LA THEORIE CHROMOSOMIQUE
En 1902, William BATESON appelle "allèles" les différentes versions d'une unité responsable d'un caractère: un individu qui possède les deux mêmes versions d'un gène est homozygote, s'il a deux versions alléliques différentes, il est hétérozygote. Il utilise en 1905 le mot "génétique" pour désigner la science qui étudie l'hérédité, c'est-à-dire la transmission des caractères et leurs variations.
Dans un article publié en 1903, "The chromosoms in heredity", SUTTON écrit:
"Les phénomènes de la division cellulaire des cellules germinales et de l'hérédité ont les mêmes caractéristiques essentielles, pureté des unités (chromosomes, caractères) et transmission indépendante de ces derniers avec comme corollaire que chacune des deux unités antagonistes est contenue dans exactement la moitié des gamètes produits."
Avec les travaux de MORGAN, qui seront vus ci-dessous, la théorie chromosomique sera reformulée de la manière suivante: les gènes (mot utilisé pour la première fois par William JOHANNSEN en 1909) sont portés par les chromosomes et occupent sur ceux-ci des emplacements fixes. Les chromosomes sont donc responsables du stockage et de la transmission du patrimoine génétique.
![]()
![]() En utilisant des couleurs différentes pour chaque couple de chromosomes, mais concordantes pour les chromosomes d'origine paternelle (couleur sombre) et pour les chromosomes d'origine maternelle (couleur claire), montrez, dans un schéma appliqué au dihybridisme, le parallélisme entre "facteurs mendéliens" et chromosomes durant la méiose.
En utilisant des couleurs différentes pour chaque couple de chromosomes, mais concordantes pour les chromosomes d'origine paternelle (couleur sombre) et pour les chromosomes d'origine maternelle (couleur claire), montrez, dans un schéma appliqué au dihybridisme, le parallélisme entre "facteurs mendéliens" et chromosomes durant la méiose.
3 Morgan: des résultats de monohybridisme différents de ceux de Mendel
En 1908, Thomas H. MORGAN est embryologiste à la Columbia University de New-York et étudie le développement d'une petite mouche des fruits, la Drosophile (Drosophila melanogaster) ou mouche du vinaigre.
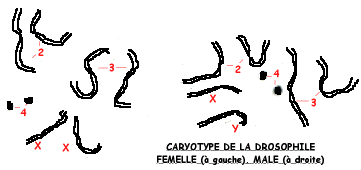
C'est un matériel de choix dans la mesure ou sa petite taille facilite l'élevage dans des flacons de verre. Chaque accouplement produit des centaines d'individus et les générations se succèdent tous les 15 jours. De plus des mutations apparaissent spontanément, dont on peut augmenter la fréquence par traitement aux rayons X. Leur garniture chromosomique est simple: 2n=8 et les mâles se distinguent des femelles par un chromosome Y et un chromosome X au lieu de deux chromosomes X.

3.1 APPARITION DE MUTANTS DANS UN ELEVAGE DE DROSOPHILES
Dans une souche dite "sauvage", il repère un mâle dont les yeux ne sont pas rouges comme ceux des autres individus, mais blancs. Il décide alors d'étudier la transmission de cette mutation appelée "white" en croisant cette mouche avec des femelles de type "sauvage", c'est-à-dire aux yeux rouges (premier croisement). Il réalise ensuite un deuxième croisement où, cette fois-ci, le caractère muté est porté par la femelle (deuxième croisement).
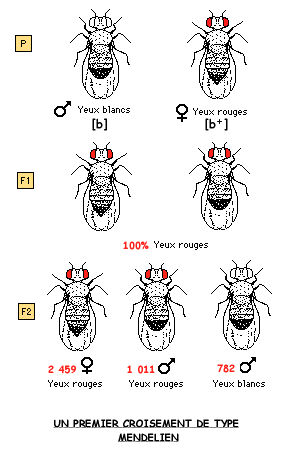
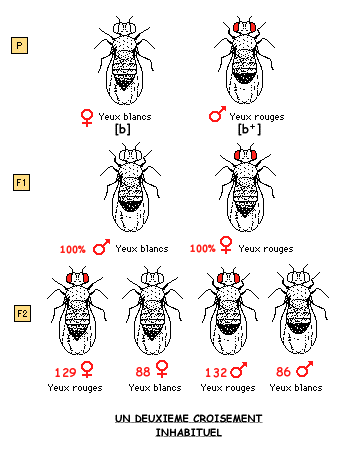
3.2 L'HEREDITE LIEE AU SEXE
Les premières observations de cellules d'insectes - comme celles de SUTTON du §2.1 - avaient révélé l'existence de chromosomes identiques appelés chromosomes homologues et un chromosome dépareillé X chez les mâles.
En 1909, une élève de MORGAN, Nettie M. STEVENS révèle un lien entre le sexe et l'existence de deux chromosomes X chez les femelles, d'un seul chromosome X et d'un chromosome Y chez les mâles.
MORGAN et BATESON, dans un premier temps, hésitent à concevoir qu'une entité telle que les chromosomes puisse engendrer les complexités de l'hérédité et du développement: ils hésitent devant une théorie moléculaire de l'hérédité.
Mais MORGAN deviendra le principal promoteur des deux théories, mendélienne et chromosomique et publiera sa propre découverte: le chromosome X de la drosophile est associé à un trait (la couleur des yeux) qui n'est pas celui du sexe.
Il affirmera un an plus tard que le chromosome X est aussi associé à des traits concernant la pigmentation du corps et la structure des ailes.
En 1911, dans un article intitulé "An attempt to analyze the constitution of the chromosomes on the basis of sex-limited inheritance in Drosophilia" MORGAN écrira donc: "ce qui est de la plus haute importance, c'est la découverte que le chromosome X ne contient pas seulement l'un des facteurs essentiels de la détermination du sexe, mais aussi tous les autres caractères qui sont liés au sexe dans l'hérédité".
![]()
![]() En utilisant les résultats des croisements observés ci-dessus, quelles sont les différences par rapport à ceux obtenus par MENDEL? Dites pourquoi les chromosomes X et Y ne peuvent pas être considérés comme des chromosomes homologues.
En utilisant les résultats des croisements observés ci-dessus, quelles sont les différences par rapport à ceux obtenus par MENDEL? Dites pourquoi les chromosomes X et Y ne peuvent pas être considérés comme des chromosomes homologues.
Pour interpréter ces résultats, MORGAN fait le raisonnement que le chromosome Y est "génétiquement vide". Sans pour cela le négliger, puisqu'il intervient dans la fécondation, vous ferez, selon la technique déployée des échiquiers du §1, l'interprétation chromosomique des croisements obtenus au §3.1
4 Morgan: des résultats de dihybridisme non conformes à la seconde loi de Mendel
4.1 DES RECOMBINAISONS INATTENDUES
Dans l'exemple ci-dessous à droite, MORGAN a croisé des drosophiles différant par des caractères dont les gènes sont portés par le chomosome X, donc des gènes liés matériellement. Pour élément de comparaison est présenté à gauche un cas de dihybridisme mendélien avec deux caractères correspondant à des gènes à ségrégation indépendante, c'est-à-dire portés par deux chromosomes homologues différents.
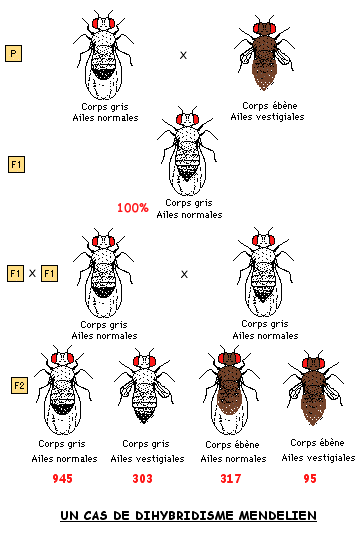
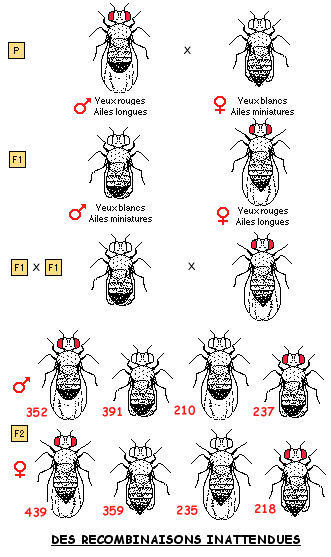
4.2 LIAISON ENTRE GENES ET BRASSAGE INTRA-CHROMOSOMIQUE
Au début de la méiose, en prophase I, on voit des chiasmas, c'est-à-dire des zones de contact entre chromatides de chromosomes homologues:
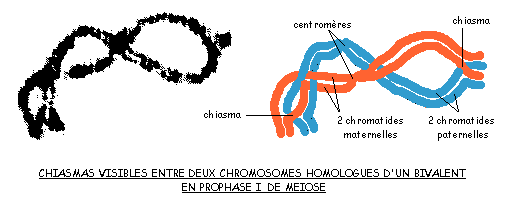
Le cytologiste belge Frans Alfons JANSSENS, en 1909, émet l'hypothèse que les chromatides pourraient se briser et se recoller en réassociant des segments homologues paternels et maternels: le résultat est une recombinaison. MORGAN propose cette hypothèse pour expliquer les réassociations entre gènes. Il nomme crossing-over cet échange physique entre segments de chromosomes.
![]()
![]() Faites l'interprétation chromosomique des résultats des deux croisements ci-dessus afin de bien comprendre la différence entre ségrégation indépendante et liaison associée au crossing-over (brassage intra-chromosomique).
Faites l'interprétation chromosomique des résultats des deux croisements ci-dessus afin de bien comprendre la différence entre ségrégation indépendante et liaison associée au crossing-over (brassage intra-chromosomique).
MORGAN crée ainsi en 1911 un nouveau concept: celui de crossing-over. Ce n'est que 20 ans plus tard que la preuve matérielle de ces échanges sera apportée par plusieurs travaux expérimentaux.
En étudiant statistiquement de nombreux croisements, MORGAN met en évidence la liaison entre gènes transmis préférentiellement ensemble: ils les appellent groupes de liaison et en définit quatre chez la drosophile.
![]() A partir de la connaissance du caryotype de la drosophile expliquez l'existence de ces quatre groupes de liaison.
A partir de la connaissance du caryotype de la drosophile expliquez l'existence de ces quatre groupes de liaison.
En définitive ...
Toutes les observations de MORGAN:
les bases de la liaison entre les caractères,
la théorie de la recombinaison entre gènes sur un seul chromosome,
la possibilité qu'un seul gène puisse influencer davantage qu'un caractère,
la nature des mutations génétiques,
consignées dans son remarquable article de 1911, vont le convaincre que non seulement la théorie chromosomique est correcte, mais aussi que MENDEL avait vu juste. De plus elles le conduiront à accepter la nature physique du gène: "la ségrégation, le point clé de tous les phénomènes mendéliens, doit se manifester dans la séparation, durant la maturation de l'oeuf et du sperme, des corps matériels (substances chimiques) contenues dans les chromosomes".
Dès 1915, MORGAN, Alfred H. STURTEVANT, Hermann J. MULLER et Calvin B. BRIDGES publieront leur traité de référence "Le mécanisme de l'hérédité mendélienne", puis, en 1917, MORGAN défendra son point de vue avec élégance dans "La théorie du gène", un article publié dans The American Naturalist.
Il écrira : "jusqu'ici, j'ai parlé du facteur génétique comme d'une unité dans le plasma germinatif dont la présence est déduite du caractère lui-même. Pourquoi, peut-on se demander, n'est-il pas plus simple de traiter des caractères eux-mêmes, comme le fit MENDEL, plutôt que d'introduire une entité imaginaire, le gène ? Il y a plusieurs raisons pour lesquelles nous avons besoin d'une conception du gène".
A ce point de son exposé, MORGAN dresse une liste des raisons pertinentes et contraignantes, et il conclut : "toutes ces preuves ont joué un rôle pour nous persuader que les gènes postulés pour l'hérédité mendélienne ont une base réelle et sont localisés sur les chromosomes".
5 Les premières cartes génétiques et la définition provisoire du gène
5.1 LA METHODE DE STURTEVANT POUR ORDONNER LES GENES
MORGAN avait remarqué que le pourcentage de recombinaison entre deux gènes liés est toujours à peu près le même et que celui-ci varie avec le couple de gènes considéré.
Il en déduit que chaque gène occupe sur un chromosome un emplacement déterminé (locus) et émet l'hypothèse d'une corrélation entre le pourcentage de crossing-over et la distance entre les gènes: ainsi le pourcentage de recombinaison, qui traduit la fréquence des crossing-over, est-elle d'autant plus grande que les gènes sont plus éloignés les uns des autres.
Un étudiant de MORGAN, A.H.STURTEVANT exploite cette idée et réalise que les distances entre gènes doivent être additives: il affirme que le pourcentage de crossing-over peut servir à déterminer la position des gènes sur le chromosome. Il construit ainsi la première carte génétique du chromosome X de Drosophila melanogaster dans les années 1910 et définit une unité de distance génétique:
1 % de crossing-over = 1 unité de distance génétique
L'unité de distance génétique sera appelée plus tard centiMorgan (cM) en hommage à MORGAN.
5.2 LES CARTES FACTORIELLES OU CARTES GENETIQUES
0n considère chez la drosophile trois couples d'allèles localisés sur le chromosome X:
|
CARACTERES |
Allèles sauvages dominants |
Allèles mutés récessifs |
|
Couleur du corps |
Corps gris j+ |
Corps jaune j |
|
Aspect des yeux |
Yeux lisses r+ |
Yeux rugueux r |
|
Longueur des ailes |
Ailes longues c+ |
Ailes coupées c |
Trois séries de croisements sont réalisés avec des femelles mutantes homozygotes pour deux gènes et des mâles sauvages, qui donnent les résultats suivants:
|
femelle [j c] x mâle [j+c+] | ||||
|
F1 |
femelle [j+c+] |
mâle [j c] | ||
|
F2 |
1 075 [j c] |
1 086 [j +c+] |
360 [j c+] |
359 [j +c] |
|
|
|
|
|
|
|
femelle [j r] x mâle [j+r+] | ||||
|
F1 |
femelle [j+r+] |
mâle [j r] | ||
|
F2 |
1 353 [j r] |
1 371 [j +r+] |
82 [j r+] |
72 [j +r] |
|
|
|
|
|
|
|
femelle [r c] x mâle [r+c+] | ||||
|
F1 |
femelle [r+c+] |
mâle [r c] | ||
|
F2 |
1 137 [r c] |
1 158 [r +c+] |
297 [r c+] |
288 [r +c] |
|
|
|
|
|
|
![]() En analysant les gènes pris deux à deux vous les situerez les uns par rapport aux autres pour en établir une carte génétique.
En analysant les gènes pris deux à deux vous les situerez les uns par rapport aux autres pour en établir une carte génétique.
Le gène, à la suite des travaux de MORGAN, apparaît ainsi comme ...
une unité de fonction, car il est indispensable à l'expression d'un caractère,
une unité de mutation, car, en subissant des changements, il donne naissance à de nouveaux allèles, donc de nouvelles formes du caractère,
une unité de recombinaison, car un allèle situé sur un chromosome, peut, à la méiose, être échangé avec un autre allèle de ce gène situé sur une chromatide du chromosome homologue.
Mais quelle est la nature chimique du gène et par quel(s) mécanisme(s) est-il à l'origine d'un caractère?