Révolution technologique des années 70
LA BIOLOGIE MOLECULAIRE ET SES APPLICATIONS
(suite)
4 Découverte des enzymes de restriction
A l'aube des années 1970, bien que les bases de l'expression des gènes soit de mieux en mieux comprises, on ne sait pas les isoler pour en faire une étude approfondie. Un pas décisif sur le plan technique va permettre de mettre au point les "outils du génie génétique".
4.1 LA BIOLOGIE DEVIENT OPERATOIRE:
Des travaux furent menés à bien en 1965 par Werner ARBER, Daniel NATHANS et Hamilton SMITH qui obtiendront le prix Nobel en 1978: ils découvrent les enzymes de restriction (endonucléases) permettant de découper l'ADN en petits segments à des endroits déterminés.
Les enzymes de restriction sont produites par les bactéries lorsqu'elles sont infectées par un bactériophage. Mais, contrairement aux DNases connues jusque là, qui coupent l'ADN au hasard, les DNases de restriction sont capables de couper l'ADN en des sites déterminés: elles reconnaissent une séquence de bases et coupent l'ADN chaque fois qu'elles la rencontrent.
Les chercheurs disposent à l'heure actuelle de plusieurs centaines de telles enzymes, véritables "ciseaux moléculaires" (Eco Ri, Hind III, Taq I...) reconnaissant des séquences de bases spécifiques et engendrant des fragments d'ADN dont les extrémités forment de véritables "bouts collants".
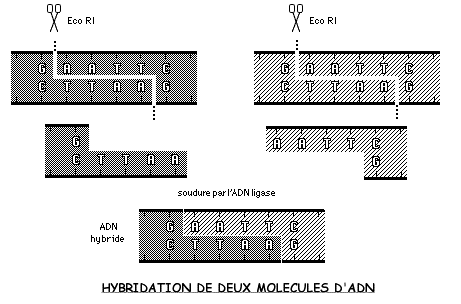 Cette propriété en fait des outils de choix permettant de découper l'ADN pratiquement où l'on veut non seulement pour isoler des gènes, mais aussi pour les insérer dans des phages ou des plasmides qui se révélèrent d'excellents vecteurs pour multiplier des gènes étrangers.
Cette propriété en fait des outils de choix permettant de découper l'ADN pratiquement où l'on veut non seulement pour isoler des gènes, mais aussi pour les insérer dans des phages ou des plasmides qui se révélèrent d'excellents vecteurs pour multiplier des gènes étrangers.
L'hybridation de deux molécules d'ADN provenant de deux espèces différentes nécessite la soudure des fragments obtenus après action des enzymes de restriction. Une enzyme, l'ADN ligase, facilite la réassociation des "bouts collants" par le biais de la complémentarité de leurs bases.
L'ARN polymérase est un complexe enzymatique qui assure plusieurs fonctions:
- reconnaissance sur l'ADN des signaux génétiques qui permettent de commencer et de terminer la synthèse d'ARNm en des sites précis,
- ouverture de la molécule d'ADN au niveau des liaisons labiles qui unissent les deux brins,
- polymérisation des nucléotides dans un ordre imposé par la complémentarité des bases des nucléotides de l'ARN et de celles des nucléotides d'une des deux chaînes transcrites de l'ADN.
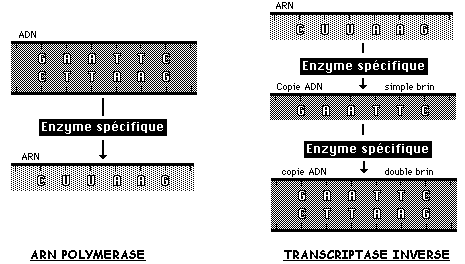 En 1970, Howard Martin TEMIN et David BALTIMORE isolent, à partir de virus, la transcriptase inverse, une enzyme capable de réaliser l'opération inverse de celle de l'ARN polymérase, qui permet d'obtenir de l'ADN à partir de l'ARN. Ils seront récompensés par le Nobel en 1975 avec Renato DULBECCO.
En 1970, Howard Martin TEMIN et David BALTIMORE isolent, à partir de virus, la transcriptase inverse, une enzyme capable de réaliser l'opération inverse de celle de l'ARN polymérase, qui permet d'obtenir de l'ADN à partir de l'ARN. Ils seront récompensés par le Nobel en 1975 avec Renato DULBECCO.
Son utilisation rend possible la synthèse d'un gène à partir de l'ARNm gouvernant l'assemblage d'une protéine. On obtient d'abord une copie d'ADN simple brin, puis à l'aide d'autres enzymes spécifiques, une copie double brin d'ADN complémentaire (ADNc).
L'hybridation ultérieure de ce gène avec un autre ADN exige la formation de nouveaux "bouts collants" à chacune de ses extrémités.
4.2 PRINCIPE D'ETABLISSEMENT D'UNE CARTE DE RESTRICTION:
![]()
![]() Cet exercice, purement méthodologique, vise à vous entraîner à tirer des conclusions par analyse logique d'un ensemble de données expérimentales. L'objectif est de rechercher les positions relatives des sites de coupures par deux enzymes de restriction sur un échantillon d'ADN de longueur 5 000 pb (paires de bases).
Cet exercice, purement méthodologique, vise à vous entraîner à tirer des conclusions par analyse logique d'un ensemble de données expérimentales. L'objectif est de rechercher les positions relatives des sites de coupures par deux enzymes de restriction sur un échantillon d'ADN de longueur 5 000 pb (paires de bases).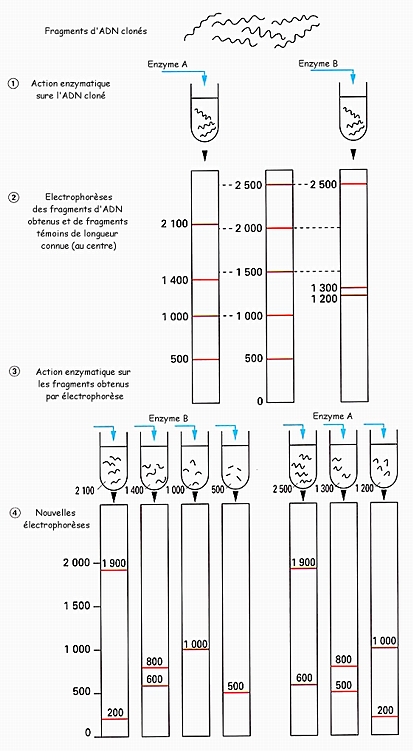 A - Les chercheurs disposent de nombreux exemplaires d'un échantillon d'ADN, obtenus par clonage de l'échantillon initial. Ce matériel est divisé en deux lots équivalents et chaque lot est soumis à l'action d'une enzyme de restriction, l'enzyme A pour un lot, l'enzyme B pour l'autre lot.
A - Les chercheurs disposent de nombreux exemplaires d'un échantillon d'ADN, obtenus par clonage de l'échantillon initial. Ce matériel est divisé en deux lots équivalents et chaque lot est soumis à l'action d'une enzyme de restriction, l'enzyme A pour un lot, l'enzyme B pour l'autre lot.
On rappelle qu'une enzyme de restriction coupe l'ADN chaque fois qu'elle rencontre un site précis, différent d'une enzyme à l'autre. L'attaque enzymatique est conduite de façon telle que tous les sites spécifiques de l'enzyme A pour un lot, de l'enzyme B pour l'autre lot, sont coupés.
Chaque lot d'ADN est alors soumis à une électrophorèse qui sépare les fragments d'ADN obtenus selon leur taille. Une électrophorèse témoin consistant à séparer des fragments d'ADN de taille connue permet de disposer d'une échelle des tailles (exprimée ici en paires de bases). Les résultats de ces premières électrophorèses sont présentés sur le document ci-contre.
1° Combien l'échantillon d'ADN initial présentait-il de sites de coupure pour l'enzyme A ? pour l'enzyme B ?
2° Montrez à l'aide de schémas simples qu'il n'est pas possible de reconstituer avec certitude la disposition relative des fragments obtenus par l'action d'une enzyme de restriction (A ou B).
B - Chaque ensemble de fragments obtenus par action de l'enzyme A est alors isolé et traité par l'enzyme B. Dans chaque cas, une nouvelle électrophorèse sépare les fragments éventuellement obtenus.
La même opération est réalisée pour les fragments obtenus au départ par l'action de l'enzyme B qui sont soumis à l'action de l'enzyme A. Les résultats de ces électrophorèses sont présentés sur le document ci-contre.
1° Comparez le résultat de l'action
- de A sur le fragment de 2 500 pb découpé par B ;
- de B sur le fragment de 2 100 pb découpé par A.
Illustrez par un schéma la conclusion se dégageant de cette comparaison.
2° Poursuivez cette comparaison méthodique de couples de fragments convenablement choisis et proposez une "carte" montrant les positions relatives des sites de coupure pour A et B sur l'échantillon d'ADN initial.
5 Le clonage et le séquençage des gènes
5.1 LE REPERAGE D'UN GENE: LA TECHNIQUE DU SOUTHERN BLOT
La combinaison de l'électrophorèse et de l'hybridation de sondes a permis de repérer la présence d'une séquence précise dans l'ADN. La technique a été mise au point par E.M. SOUTHERN (1975):
l'ADN est traité par une enzyme de restriction: plusieurs milliers de fragments de tailles différentes sont ainsi produits.
le mélange est disposé dans un puits d'une plaque de gel d'agarose et soumis à une électrophorèse qui provoque la migration des fragments, d'autant plus vite qu'ils sont petits.
après dénaturation de l'ADN (séparation des brins complémentaires), la plaque est mise au contact d'un filtre de nitrocellulose qui recueille une empreinte ("blot") du gel.
le filtre est immergé dans une solution contenant une sonde moléculaire radioactive (fragment d'ADN reconnaissant une séquence du génome) pendant quelques heures à 65°C: celle-ci se fixe sur les fragments complémentaires. Le filtre est ensuite rincé pour éliminer les sondes non fixées.
on procède à la révélation de l'emplacement de la sonde et donc des fragments complémentaires par une technique appropriée : autoradiographie pour les sondes radioactives, photographie pour les sondes fluorescentes, réactions enzymatiques pour les ligands reconnus par des protéines spécifiques.
Cette technique est très sensible et permet de reconnaître un fragment sur les milliers au départ. Elle permet ainsi de faire la recherche simultanée chez plusieurs individus et de procurer une carte génétique d'une grande spécificité et fiabilité, par construction d'une carte de restriction. L'analyse est appelée polymorphisme de longueurs de fragments de restriction (RFLP: "restriction fragment length polymorphism).
5.2 LE CLONAGE D'UN GENE:
Les enzymes de restriction ont ouvert la voie au clonage des gènes à partir de 1972, dans le but de les reproduire.
Supposons que l'ADN d'une cellule humaine ait été découpé et ses fragments transférés chez E.coli: on dispose ainsi d'une "banque génomique" constituée de milliers de clones bactériens différents, chacun ayant multiplié un fragment du génome humain. La technique du Southern blot permet de repérer le clone ayant incorporé le gène recherché.
Une fois le gène isolé, il est introduit dans un vecteur qui permet son amplification et son stockage dans une cellule vivante, généralement une bactérie.
5.2.1 Les plasmides bactériens:
De nombreuses bactéries contiennent un ou plusieurs plasmides: la plupart sont des molécules d'ADN circulaires double-brin, beaucoup plus petite qu'un chromosome de 3 à 10 kb (milliers de paires de bases).
Ce sont des unités de réplication autonome vis-à-vis du chromosome bactérien, mais qui nécessite la machinerie de la cellule pour leur réplication.
Ils ne sont pas indispensables au métabolisme de la cellule dans des conditions normales de croissance.
Les plasmides peuvent coder pour diverses fonctions:
Certains codent pour des enzymes inactivant des antibiotiques: de tels plasmides R (portant un gène de résistance) rendent habituellement la cellule hôte résistante à un antibiotique.
Le facteur F est un plasmide particulier jouant un rôle dans la sexualité des bactéries. Il peut-être soit absent des cellules (F-) soit présent sous deux états possibles : autonome et indépendant du chromosome (bactéries F+) ou intégré dans le chromosome bactérien (bactéries HFR).
Certains plasmides possèdent l'information nécessaire pour se transférer eux-mêmes d'une cellule à l'autre par conjugaison.
Les plasmides sont largement utilisés dans les techniques de l'ADN recombinant. Un plasmide recombinant (manipulé génétiquement) est souvent désigné par le préfixe "p" (par exemple pBR329).
5.2.2 Principe de l'utilisation des plasmides
Les plasmides sont utilisés comme vecteur de clonage
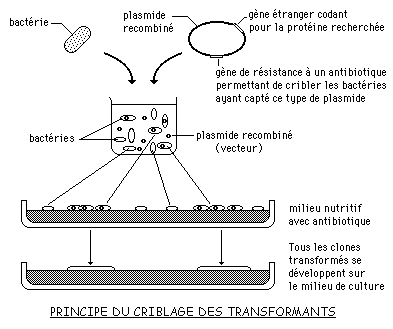 Par des expériences de recombinaison in vitro, on sait intégrer dans un plasmide un fragment d'ADN provenant d'une autre source: on obtient ainsi un plasmide recombiné ou plasmide chimère.
Par des expériences de recombinaison in vitro, on sait intégrer dans un plasmide un fragment d'ADN provenant d'une autre source: on obtient ainsi un plasmide recombiné ou plasmide chimère.
On peut faire pénétrer ces molécules recombinées dans des cellules d'Escherichia coli dépourvues de plasmide (transformation bactérienne).
On peut repérer - on dit cribler - les cellules qui ont reçu une molécule de plasmide grâce aux gènes du plasmide qui sont sélectionnables (résistance à un antibiotique par exemple). Parmi ces cellules, on pourra reconnaître (ou sélectionner) celles qui portent un plasmide ayant intégré un fragment particulier de l'ADN étranger.
L'ensemble des descendants d'une telle cellule constitue un clone. On dit que l'on a cloné un fragment d'ADN.
Ce fragment sera beaucoup plus facile à obtenir, à purifier, et à étudier dans un clone bactérien que dans son contexte d'origine (noyau d'une cellule de mammifère par exemple).
Les plasmides sont utilisés comme vecteur d'expression
Une fois un fragment d'ADN cloné dans un plasmide on peut réussir à faire exprimer les gènes qu'il porte dans les cellules d'Escherichia coli. Cette expression nécessite la transcription de ce fragment et la traduction du messager. On peut ainsi faire fabriquer à Escherichia coli des protéines étrangères (par exemple l'insuline humaine, l'hormone de croissance humaine).
Les plasmides sont utilisés comme vecteur pour la transformation de cellules eucaryotes
On sait depuis quelques années introduire et faire exprimer de l'ADN dans des cellules eucaryotes (levure, cellules de Mammifères en culture, cellules résultant des premières divisions de l'oeuf d'un mammifère par exemple). On peut ainsi, après un clonage dans Escherichia coli, réintroduire le gène cloné dans des cellules eucaryotes et le faire exprimer.
5.3 LE SEQUENCAGE DE L'ADN:
Le séquençage de l'ADN consiste à déterminer l'ordre des nucléotides sur la molécule d'ADN. C'est le moyen le plus précis pour rechercher la présence de mutations ponctuelles dans un gène.
La méthode utilisée aujourd'hui, proposée par F. SANGER en 1977, qui a obtenu le prix Nobel de chimie en 1980, est devenue aujourd'hui une technique rapide et fiable utilisée fréquemment dans le diagnostic des maladies héréditaires.
6 La nouvelle conception du gène
6.1 LE MORCELAGE DU GENE DES EUCARYOTES
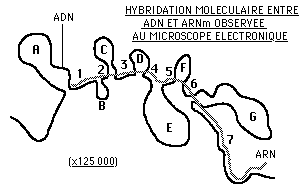 Le gène de l'ovalbumine: les travaux de P.CHAMBON (1977)
Le gène de l'ovalbumine: les travaux de P.CHAMBON (1977)
Le document ci-contre traduit le résultat d'une expérience d'hybridation au cours de laquelle le brin d'ADN codant pour l'ovalbumine a été mis en présence de l'ARN messager qui, dans le cytoplasme, dirige la synthèse de cette protéine. Les nucléotides appartenant aux séquences complémentaires de l'ADN et de l'ARN vont se reconnaître et s'accoler par leurs bases.
![]()
![]() Décrivez ce que vous observez et tirez-en une conclusion quant au mode d'expression d'un gène d'eucaryote par rapport à celui d'un procaryote. Proposez un schéma pour illustrer cette différence.
Décrivez ce que vous observez et tirez-en une conclusion quant au mode d'expression d'un gène d'eucaryote par rapport à celui d'un procaryote. Proposez un schéma pour illustrer cette différence.
On appelle introns les séquences d'ADN transcrites, puis excisées lors de la maturation de l'ARNpm (ARN prémessager). Les séquences d'ADN transcrites en ARNpm et conservées dans l'ARNm après excision sont appelées exons.
La découverte d'introns et d'exons dans les gènes d'eucaryotes introduit ainsi un nouveau concept de gène: celui de gène morcelé, qui va conduire à réviser le dogme "un gène -> une protéine".
6.2 CONSEQUENCE DU MORCELLEMENT DES GENES
Les exons d'un ARN pré-messager ne sont pas toujours épissés de la même manière: ceci va donner naissance à des ARN messagers différents, donc à des protéines différentes (épissage alternatif). Un même gène peut ainsi gouverner la synthèse de protéines différentes: c'est le cas du gène de la calcitonine.
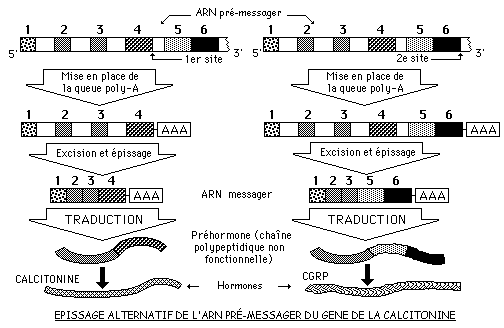
L'hormone CALCITONINE est une hormone produite et sécrétée par des cellules thyroïdiennes.
L'hormone CGRP (Calcitonine Gene Related Protéine) est un neuropeptide produit par certaines cellules de l'hypophyse.
Dans des cellules thyroïdiennes, l'ARNpm est découpé et polyadénylé au 1er site. L'excision de 3 introns et l'épissage des 4 exons restants produisent l'ARNm codant pour un précurseur de la calcitonine.
Dans des cellules hypophysaires, le même ARNpm est découpé au 2e site. Après excision de 4 introns, de l'exon 4 et épissage de 5 exons, l'ARNm obtenu code pour le précurseur du CGRP.
On estime que plus de 40% des gènes humains sont l'objet d'un épissage alternatif: ceci confère au génome une possibilité de synthèse de protéines bien supérieure au nombre de gènes. C'est ce que nous constaterons pour expliquer la grande variété des anticorps que l'organisme est capable de produire, eu égard aux 30 à 40 000 gènes du génome humain.
7 Les OGM
7.1 LA TRANSGENESE
Le génie génétique permet d'introduire dans une cellule un gène qu'elle ne détient pas. Dans son environnement antérieur ce gène s'exprime habituellement, c'est-à-dire qu'il code pour la synthèse d'une protéine.
Il est possible également de supprimer ou de modifier l'expression d'un gène déjà présent dans le génome d'une cellule.
Le génie génétique peut s'appliquer aux végétaux, aux animaux, à l'homme, aux micro-organismes.
Une plante transgénique est une plante dont le génome a été modifié par l'introduction d'un gène qui peut provenir d'une autre plante, d'une bactérie ou de tout autre organisme. Ce gène peut coder une enzyme qui intervient dans la maturation des fruits, une substance qui bloque la multiplication d'un virus, une nouvelle protéine, par exemple un composé toxique pour les insectes ravageurs, ... C'est ce dernier exemple que nous exposerons.
7.1.1 Qu'entend-on par OGM?
Il est nécessaire de faire cette précision afin de conduire le débat que nous pourrons avoir pour répondre aux inquiétudes que ces OGM suscitent.
Un OGM (Organisme Génétiquement Modifié) est un organisme vivant dont le génome a été modifié par génie génétique. Toutes les cellules de cet organisme possèdent le gène étranger: cette modification génétique est donc transmissible à sa descendance par le biais de toutes ses parties capables de donner une nouvelle plante (fruits, graines, organes de reproduction végétative).
Les produits dérivés des OGM, n'ayant aucune capacité de reproduction, ne sont pas des OGM, même s'ils peuvent éventuellement contenir le gène introduit ou la protéine codée par ce gène.
Ainsi, des plants de colza transgénique résistant à un champignon pathogène sont des OGM, leurs graines aussi.
En revanche, l'huile de colza destinée à l'alimentation de l'homme ou le tourteau de colza destiné à l'alimentation animale, ne sont pas des OGM, mais des produits dérivés d'un OGM.
Compte tenu des procédés de fabrication, il est très probable que la plupart des huiles raffinées que nous consommons ne contiennnent pas de protéines ou d'ADN.
Le tourteau, obtenu à partir des résidus de fabrication de l'huile, contient des protéines et de l'ADN. Il peut donc contenir le gène de résistance au champignon pathogène et la protéine codée par ce gène.
7.1.2 Comment obtient-on une plante transgénique?
Après avoir repéré un caractère intéressant dans un autre organisme vivant (plante, champignon, bactérie...) et identifié la protéine responsable de ce caractère, on identifie et on isole le gène codant cette protéine.
On réalise une "construction génique" qui contient le gène d'intérêt et des séquences d'ADN (promoteur, terminateur) indispensables à son fonctionnement dans le génome d'une cellule végétale. Ces séquences sont impliquées dans la régulation de l'expression du gène. Elles permettent de cibler le lieu d'expression du gène dans la plante (graines, racines, feuilles...), voire de faire en sorte qu'il ne s'exprime qu'au moment nécessaire, lors de l'attaque d'un insecte ou de l'infection par un virus par exemple. Cette construction contient éventuellement un gène marqueur de repérage des plantes transgéniques (par exemple un gène de résistance à un antibiotique: voir ci-dessus § 5.1.2). Cette construction génique est ensuite insérée dans un plasmide bactérien pour être multipliée.
On introduit la construction génique dans le génome de la cellule végétale par deux méthodes:
o transfert biologique: au moyen d'un vecteur, la bactérie du sol Agrobacterium tumefaciens, qui transfère naturellement une partie de son ADN (auquel on a donc ajouté la construction génique à intégrer) dans le génome des plantes.
o transfert mécanique: les constructions géniques, portées par des microbilles de tungstène, sont projetées dans la cellule végétale.
On sélectionne les cellules exprimant le gène ajouté.
On régénère des plantes entières à partir de ces cellules. Ces plantes sont testées en serre puis en champ afin de vérifier la conformité de leur développement, la stabilité de l'expression du gène ajouté, sa transmission à la descendance.
7.2 UN EXEMPLE D'ORGANISME VEGETAL GENETIQUEMENT MODIFIE POUR LA RESISTANCE AUX INSECTES
A la suite de travaux expérimentaux, on sait que certains gènes de la bactérie Bacillus thuringiensis (Bt) transférés dans une plante lui font produire des toxines efficaces contre les insectes parasites (notamment les larves de papillons). Ces toxines ne sont pas dangereuses pour les Mammifères. Des constructions d'ADN contenant ces gènes ont été greffées à différentes plantes (coton, tomate, pomme de terre, maïs).
Donner aux plantes la possibilité de se défendre contre les insectes présente de nombreux avantages sur l'utilisation des insecticides: il n'y a plus de risque de "lessivage" de l'insecticide par les eaux de ruissellement, seuls les insectes qui attaquent la plante sont tués et la plante est protégée.
La lutte contre la Pyrale du Maïs
On trouvera dans la base de données encyclopédique HYPPZ, du serveur de l' INRA, des renseignements à propos du Maïs et de la Pyrale du Maïs.
Le maïs est attaqué par les chenilles de la pyrale. Toutes les parties de l'appareil aérien de la plante peuvent l'être, mais les dégâts les plus importants du point de vue économique se font au niveau des épis.
Les larves peuvent entrer dans l'épi et causer des dégâts importants en rongeant les grains qui sont alors impropres à la vente. De plus, un champ de maïs infesté par la pyrale peut attirer des oiseaux qui viennent chercher des insectes et se mettent ensuite à picorer les épis.
![]() En utilisant les données graphiques, textuelles du document "Maïs résistant à la Pyrale", vous essaierez de:
En utilisant les données graphiques, textuelles du document "Maïs résistant à la Pyrale", vous essaierez de:
comprendre la nécessité pour les agriculteurs de lutter contre la Pyrale du Maïs,
dégager les principes et les intérêts d'une lutte biologique contre ce parasite,
comprendre sur quoi repose la polémique actuelle sur l'utilisation de la transgenèse
7.3 PRODUCTION DE MOLECULES PHARMACOLOGIQUES
Production d'hormone de croissance
Sécrétée tout au long de la vie, l 'hormone de croissance (GH, Growth Hormone ou STH, Somatotrophic hormone) est produite par l'hypophyse humaine. Elle stimule la croissance de l'enfant et joue un rôle important dans le métabolisme des protéines, des lipides et certains glucides chez l'adulte.
Des quantités trop faibles de GH dans l'organisme d'un jeune enfant conduit au nanisme.
Elle a été isolée pour la première fois en 1956 et sa structure a été identifiée en 1972. L'hormone de croissance bovine est sans effet et jusque vers le milieu des années 80, la seule source d'hormone de croissance était l'hypophyse humaine prélevée post-mortem.
Les problèmes se posant au corps médical sont un approvisionnement limite d'hypophyse et, dans des cas rares, la transmission de maladies (comme la maladie de Creuzfeldt-Jacob qui entraîne une dégénérescence du cerveau).
Les documents abordés par le § 7 vous ont permis d'appréhender, par deux exemples, les risques (pour la santé, pour l'environnement), les avantages et les inconvénients des OGM. Vous discuterez de l'intérêt et des obstacles à franchir pour que ces OGM deviennent scientifiquement et éthiquement acceptables.
Pour forger les points de vue et étayer l'argumentation, citons le site du ministère de l'agriculture: http://www.agriculture.gouv.fr/OGM/ogm.htm